Avaluació de les asimetries de potència, cinemàtica i oxigenació muscular entre les cames durant una carrera incremental de 400 m en triatletes
Jordi Montraveta
Ignacio Fernández-Jarillo
Xavier Iglesias

Diego Chaverri
*Correspondència: Diego Chaverri dchaverri@gencat.cat
Citació
Montraveta, J., Fernández-Jarillo, I., Iglesias, X., & Chaverri, D. (2026). Assessing power, kinematic, and muscle oxygenation asymmetries between legs during incremental 400-m track running in triathletes. Apunts. Educación Física y Deportes, 165, 70-81. https://doi.org/10.5672/apunts.2014-0983.es.2026.165.07
Resum
Córrer sobre una pista de 400 m pot induir asimetries en la potència desenvolupada (PD), la cinemàtica o la saturació muscular d’oxigen (SmO₂). L’objectiu principal d’aquest estudi era determinar si córrer sobre una pista de 400 m indueix diferències mesurables en la potència desenvolupada, la cinemàtica i la saturació muscular d’oxigen entre la cama interior i l’exterior. L’objectiu secundari era avaluar la magnitud de les asimetries en la potència desenvolupada, la cinemàtica i la saturació muscular d’oxigen en triatletes durant la cursa en pista. Les dades cinemàtiques i de PD es van recollir per mitjà de sensors Stryd col·locats en les dues sabatilles, mentre que les dades d’SmO₂ es van obtenir per mitjà de dispositius d’espectroscòpia d’infrarojos propers (NIRS, per les seves sigles en anglès) col·locats sobre el múscul vast lateral (VL) de les dues cames. Encara que l’SmO₂, la PD i els paràmetres cinemàtics van mostrar canvis corresponents a l’augment de la velocitat durant el test de VAM-EVAL, un protocol de cursa incremental en el qual la velocitat augmenta a raó de 0.5 km·h-1 cada minut fins a l’esgotament, l’ANOVA de mesures repetides va revelar efectes principals significatius de la cama (interior vs. exterior) en la potència desenvolupada (PD), la longitud de gambada (LG), el temps de contacte amb el terra (TCT) i l’oscil·lació vertical (OV). Tanmateix, les anàlisis post hoc només van identificar diferències entre extremitats significatives en la PD al 80 % de la velocitat aeròbica màxima (VAM), i en l’LG al 70 % i al 80 % de la VAM. Quant a les asimetries, l’ANOVA de mesures repetides no va mostrar diferències significatives entre intensitats, excepte en l’SmO₂, amb canvis significatius observats entre el 60 % i el 90 % i entre el 70 % i el 90 % de la VAM. Aquestes observacions posen en relleu el potencial de les asimetries d’SmO₂ mesurades mitjançant NIRS per detectar de forma no invasiva diferències en la utilització d’oxigen entre una cama i l’altra, especialment a intensitats elevades (90 % de la VAM). Aquesta tecnologia podria ajudar esportistes i entrenadors a identificar desequilibris en l’aportació i la utilització d’oxigen entre cames, factor crític per optimitzar el rendiment muscular.
Introducció
Córrer sobre una pista de 400 m és fonamental per a l’entrenament i l’avaluació del rendiment, ja que proporciona un entorn controlat per valorar la resistència i l’eficiència cinemàtica (Léger i Boucher, 1980). Aquestes pistes inclouen dues rectes (d’aproximadament 84.39 m cadascuna) i dues corbes amb un radi d’uns 36.5 m (World Athletics, 2019). Les corbes plantegen desafiaments biomecànics específics, ja que obliguen els corredors a gestionar girs continus cap a l’esquerra. Aquest moviment asimètric imposa diferents demandes mecàniques i fisiològiques sobre la cama interior i l’exterior a causa de les variacions en la càrrega, amb possibles conseqüències per al rendiment i el risc de lesions (Alt et al., 2015). En suport d’aquesta perspectiva, Gilgen-Ammann et al. (2017) van observar que l’asimetria de la marxa era significativament més gran en corredors amb antecedents de lesió que en corredors sense lesions, la qual cosa suggereix que les asimetries persistents podrien reflectir vulnerabilitats subjacents. Això planteja la qüestió de si les asimetries preexistents podrien ser la causa d’aquestes lesions (Knapik et al., 1991; Vasquez-Bonilla et al., 2022, 2023).
Per comprendre millor l’origen d’aquestes asimetries, és essencial analitzar la manera en què la cursa en corba afecta determinades variables cinemàtiques com són la cadència (CAD), la longitud de gambada (LG) i el temps de contacte amb el terra (TCT). En resposta a les forces centrífugues generades durant el gir, els esportistes solen presentar una LG més curta i un TCT més llarg en la cama interior que en l’exterior, especialment a velocitats de cursa més altes. La cama interior actua com a estabilitzadora sota forces compressives més elevades, mentre que l’exterior proporciona una propulsió més gran per donar resposta a la demanda biomecànica de la corba (Chang i Kram, 2007). En l’estudi de Chang i Kram (2007), es van observar asimetries entre la cama interior i l’exterior durant esprints en una corba de 6 m de radi. La cama exterior va mostrar una LG significativament més gran que la interior (1.70 ± 0.10 m en front de 1.53 ± 0.02 m; p < .05). Encara que també es van observar diferències en la freqüència de pas (FP) (3.88 ± 0.13 passos s⁻¹ en front de 3.56 ± 0.12 passos s⁻¹) i el TCT (0.203 ± 0.008 s en front de 0.190 ± 0.006 s), aquestes diferències van mancar de significació estadística. Hamill et al. (1987) van investigar la manera en què els corredors ajustaven la funció de les extremitats inferiors en recórrer les corbes d’una pista de 400 m. El seu estudi va revelar asimetries significatives entre les extremitats dreta i esquerra durant la cursa en corba, especialment en la fase de presa del contacte del peu. Concretament, es van observar diferències en els angles de presa de contacte, els angles màxims de pronació i les forces de reacció del terra mediolaterals. Aquestes observacions suggereixen que les transicions entre les seccions corbes i rectes de la pista obliguen els corredors a adaptar dinàmicament els seus patrons de cursa. Els girs continus cap a l’esquerra en les corbes imposen demandes mecàniques i fisiològiques diferents a cada cama, cosa que pot afectar variables com l’oscil·lació vertical (OV) i el CAD (Hamill et al. 1987).
Amb el temps, aquestes càrregues mecàniques asimètriques podrien contribuir al desenvolupament d’asimetries entre extremitats, especialment en esportistes que entrenen amb freqüència en pistes amb corbes. Aquest concepte fa referència a desequilibris de rendiment entre les extremitats oposades del cos (Bishop et al., 2017, 2022; Fox et al., 2023). Si bé les asimetries entre extremitats s’han estudiat tradicionalment en esports multidireccionals que impliquen accions explosives i canvis ràpids de direcció (Bishop et al., 2017; Loturco et al., 2019), alguns estudis recents també han analitzat aquests desequilibris en disciplines lineals i de resistència com la cursa i el triatló (Helme et al., 2021; D’Hondt et al., 2024; Jacques et al., 2021). Aquestes observacions suggereixen que les asimetries són rellevants no només en els esports d’equip, sinó també en activitats caracteritzades per un moviment continu i repetitiu. Les asimetries en la distribució de la massa magra, la força muscular o la potència desenvolupada (PD) s’han relacionat amb problemes de rendiment i amb un risc més gran de lesions (Bell et al., 2014; Knapik et al., 1991).
Les revisions sistemàtiques han subratllat la complexa relació entre l’asimetria entre extremitats i el rendiment esportiu. Per exemple, Bishop et al. (2017) van concloure que les asimetries entre extremitats observades en un ampli ventall de tasques solen tenir un efecte negatiu sobre el rendiment físic. Tanmateix, les seves conclusions van posar en relleu una variabilitat considerable, ja que alguns estudis van notificar una associació mínima o nul·la entre l’asimetria i els resultats de rendiment. De la mateixa manera, D’Hondt et al. (2024) van assenyalar una elevada heterogeneïtat metodològica en els estudis sobre asimetria entre extremitats i cursa de resistència, fet que dificulta extreure conclusions definitives. Malgrat que alguns indicadors d’asimetria, com les asimetries superiors al 10 % en el salt amb contramoviment unilateral (SCML) o les diferències de força entre extremitats, poden afectar negativament al rendiment en cursa (Bishop et al. 2017), les dades científiques no sempre són coherents, la qual cosa subratlla la necessitat d’estudis de qualitat per determinar llindars i conseqüències pràctiques per a l’entrenament.
En el triatló, les asimetries cinemàtiques es podrien veure més influïdes per les demandes mecàniques prolongades i repetitives del tram amb bicicleta que per la mateixa transició. La fatiga acumulada durant el tram amb bicicleta pot alterar el control neuromuscular durant la cursa posterior, la qual cosa podria augmentar les asimetries entre extremitats (Millet i Vleck, 2000). Heiden i Burnett (2003) van demostrar que els patrons d’activació muscular a les cames durant la cursa s’alteren significativament després del tram amb bicicleta, amb canvis notables a músculs importants de les extremitats inferiors com el bíceps femoral i el vast lateral. De la mateixa manera, Connick i Li (2015) van notificar un augment de la variabilitat del temps de gambada i una reducció de la longitud de gambada durant la cursa posterior al tram amb bicicleta, la qual cosa indica una alteració del control locomotor probablement vinculada a la fatiga prèvia. Olcina et al. (2019) van observar una disminució de la longitud de la gambada després del tram amb bicicleta, fet que apunta a una alteració de l’activació neuromuscular com a factor contribuent.
A més de les variables cinemàtiques i d’activació muscular, els paràmetres de fisiologia muscular també són rellevants per comprendre millor les asimetries entre extremitats. La saturació muscular d’oxigen (SmO₂) proporciona una informació valuosa per avaluar possibles diferències entre extremitats. Una disminució de l’SmO₂ reflecteix un desequilibri més gran entre l’aportació i la utilització d’oxigen, fet que indica una major extracció muscular d’oxigen durant l’exercici (van der Zwaard et al., 2016). Mitjançant espectroscòpia d’infrarojos propers (NIRS), els investigadors han pogut avaluar si existeix variabilitat o asimetries en la utilització d’oxigen entre una cama i l’altra durant activitats com ara el ciclisme (Sendra-Pérez et al. 2025); Skotzke et al., 2024). Olcina et al. (2019) van analitzar l’efecte del tram amb bicicleta previ sobre el rendiment en la cursa, la longitud de la gambada i la saturació muscular d’oxigen (SmO₂) en triatletes, i els seus resultats van indicar que el tram amb bicicleta d’alta intensitat abans de la cursa pot perjudicar el rendiment en reduir la longitud de gambada i limitar la utilització perifèrica d’oxigen a músculs que presenten una concentració elevada d’SmO₂. Aquestes observacions suggereixen que la fatiga prèvia induïda per l’esforç amb bicicleta no només afecta l’eficiència biomecànica, sinó que també genera desequilibris localitzats d’oxigenació, la qual cosa podria agreujar les asimetries musculars durant el tram de cursa posterior. S’han notificat observacions similars en altres esports com el futbol i el rugbi, en els quals les asimetries d’SmO₂ es correlacionen amb una menor eficiència en el rendiment i un risc més gran de lesions, (Vasquez-Bonilla et al., 2022, 2023).
Actualment, la tecnologia portàtil permet fer un seguiment en temps real de les variables cinemàtiques, la PD i l’SmO₂ per avaluar les asimetries. Dispositius com el sensor Stryd proporcionen dades detallades sobre la PD, l’LG, el TCT i l’OV, que faciliten l’anàlisi del rendiment, (van Rassel et al., el 2023). Aquestes eines ajuden esportistes i entrenadors a identificar diferències entre extremitats en la producció mecànica i l’SmO₂, la qual cosa permet orientar l’entrenament per optimitzar el rendiment i reduir el risc de lesions. En estudis previs, s’ha demostrat la seva utilitat en aquest context. Per exemple, Bini i Hume (2015) van trobar asimetries notables en la força aplicada al pedal durant proves contrarellotge de ciclisme, mentre que Yanci (2014) va notificar asimetries de PD entre la cama dominant i la no dominant en esportistes de resistència. Ambdós estudis suggereixen que abordar aquests desequilibris podria millorar el rendiment i reduir el risc de lesions.
Sobre aquesta base, l’asimetria entre extremitats es defineix com el desequilibri o diferència entre les extremitats inferiors que pot manifestar-se en dimensions funcionals, cinemàtiques o cinètiques. En corredors de resistència, la investigació ha notificat asimetries funcionals d’aproximadament el 16-17 % i diferències biomecàniques que oscil·len entre el 3 % i el 54 %, segons la tasca i l’indicador que es faci servir (D’Hondt et al., 2024). Si bé les dades que quantifiquen específicament la seva prevalença general en triatletes continuen sent escasses a causa de la falta d’estudis previs en aquesta població, s’han documentat diferències laterals significatives en característiques neuromusculars. En concret, s’ha demostrat que córrer després de pedalar altera els patrons d’activació muscular (per exemple, redueix l’activació del soli en una extremitat), malgrat no observar-se diferències en la cinètica o la cinemàtica externes (Jacques et al., 2021). Ateses les demandes biomecàniques de la cursa en pista i la presència documentada d’asimetries entre extremitats en esportistes de resistència i triatletes, el present estudi es va plantejar els següents objectius:
Analitzar si córrer sobre una pista de 400 m indueix diferències mesurables en la potència, la cinemàtica i la saturació muscular d’oxigen entre la cama interior i l’exterior.
Avaluar la magnitud de les asimetries de potència, cinemàtica i saturació muscular d’oxigen en triatletes durant la cursa en pista.
Materials i mètodes
Participants
Catorze triatletes homes de nivell nacional van participar voluntàriament en aquest estudi (Taula 1). Els criteris d’inclusió van ser estar federats i competir activament en proves nacionals de triatló. Els criteris d’exclusió van ser patir algun trastorn cardíac, presentar una lesió en aquell moment o haver-la patit en els dos mesos anteriors. A més, es va excloure les persones amb un gruix del teixit adipós (GTA) superior a 7 mm per minimitzar la interferència amb la qualitat del senyal NIRS, ja que un GTA més gran pot atenuar la llum infraroja propera i afectar la precisió de les lectures d’SmO₂ (McManus et al., 2018). El GTA es va calcular com 0.5 × el gruix mitjà del plec cutani. Tots els participants van donar el seu consentiment informat per escrit, i el protocol de l’estudi va ser aprovat pel Comitè d’Ètica de Recerca Clínica de l’Administració Esportiva Catalana (026/CEICGC/2023).

Materials
Els paràmetres cinemàtics i de PD es van mesurar mitjançant el sensor Stryd (Stryd Inc., Boulder, CO, EE. UU.), que s’ha validat prèviament per a l’estimació de la potència en cursa en condicions controlades (Cerezuela-Espejo et al., 2021). Els sensors Stryd es van fixar als cordons de cada sabatilla, col·locats en posició central sobre el dors del peu, d’acord amb les instruccions del fabricant. El dispositiu va obtenir mostres i va exportar les dades processades a 1 Hz, mentre que les dades inercials internes es van registrar a freqüències més altes (fins a ~100 Hz). L’SmO2 es va mesurar amb un dispositiu NIRS d’ona contínua (MOXY, Hutchinson, Minnesota, Estats Units) utilitzant quatre longituds d’ona (680, 720, 760 i 800 nm). L’SmO2 es va mesurar en forma de percentatge aplicant la següent equació: SmO2 = [oxihemoglobina (O2Hb) / (oxihemoglobina (O2Hb) + desoxihemoglobina (HHb))] * 100, on O₂Hb representa l’oxihemoglobina i HHb representa la desoxihemoglobina. El dispositiu MOXY és un sistema NIRS portàtil i no invasiu dissenyat per a avaluacions de camp de l’oxigenació muscular. La seva validesa i fiabilitat per mesurar l’SmO₂ durant l’exercici s’han demostrat prèviament (Feldmann et al., 2019). El dispositiu NIRS es va col·locar sobre la zona central del múscul vast lateral (VL) a ambdues cames, a mig camí entre el trocànter major i l’epicòndil femoral lateral (McManus et al., 2018). Els sensors es van fixar amb cinta adhesiva (Hypafix; BSN Medical, Hamburg, Alemanya). La freqüència de mostratge es va deixar en la manera predeterminada del dispositiu: les dades s’actualitzaven cada dos segons i se suavitzaven prenent com a referència els 10 segons anteriors.
Procediments
La col·locació dels sensors Moxy es va determinar amb una cinta mètrica i un retolador. A més, el gruix del plec cutani es va mesurar al lloc on es va col·locar el dispositiu NIRS mitjançant un adipòmetre Harpenden (precisió: 0.20 mm). A continuació, es van fixar els sensors Stryd a les dues sabatilles.
Tots els participants van fer un test de VAM-EVAL (García i Secchi, 2013) sobre una pista de 400 m.Segons el protocol original, no es va fer un escalfament estandarditzat abans de la prova, ja que la baixa velocitat inicial de cursa proporcionava un escalfament progressiu.La velocitat de cursa es va controlar mitjançant un senyal acústic. La velocitat inicial es va establir en 8.5 km·h⁻¹ i va augmentar a raó de 0.5 km·h⁻¹ per minut. Es van col·locar cons cada 20 m per ajudar els participants a ajustar el seu ritme als senyals acústics. La prova va finalitzar quan els participants es van aturar per esgotament o no van assolir el con marcat en dues ocasions consecutives. La velocitat aeròbica màxima (VAM) es va definir com la velocitat assolida durant l’última etapa completada; no es van tenir en compte les etapes incompletes (García i Secchi, 2013).
Les sessions de prova es van fer en una pista d’atletisme a l’aire lliure a Barcelona entre el 27 de febrer i el 23 d’abril de 2024, en condicions ambientals mitjanes de 21.1 ± 5.7 °C i 74.6 ± 16.6 % d’humitat relativa. Es va indicar als participants que s’abstinguessin de fer entrenaments d’alt volum o alta intensitat durant les 24 hores prèvies a la prova.
Anàlisi estadística
Les dades cinemàtiques i d’SmO₂ es van filtrar cada 2 segons abans de l’anàlisi i es van alinear temporalment amb el temps zero (t = 0) per a la seva sincronització. El temps zero es va definir com l’inici del test de VAM-EVAL, identificat pel primer augment de la velocitat de cursa indicat pel senyal acústic, i es va sincronitzar entre dispositius utilitzant les marques temporals internes dels sistemes de registre. Es van calcular dades estadístiques descriptives (mitjana ± desviació típica [DT]) per a totes les variables.
Els nivells d’intensitat (60 %, 70 %, 80 % i 90 % de la velocitat aeròbica màxima [VAM]) es van determinar a partir dels resultats del test de VAM-EVAL. Els valors mitjans d’ambdues cames es van obtenir a partir dels 20 segons centrals de cada interval d’un minut en cada nivell d’intensitat.
La normalitat es va avaluar mitjançant la prova de Shapiro-Wilk. Amb l’excepció d’alguns índexs d’asimetria (PD ASIM, CAD ASIM, OV ASIM, TCT ASIM i LG ASIM), totes les variables van complir el supòsit de normalitat. En conseqüència, es va fer una anàlisi de la variància (ANOVA) de mesures repetides de dos factors per comparar les diferències entre cames (cama interior davant cama exterior) i nivells d’intensitat (60 % , 70 %, 80 % i 90 % de la VAM) en la PD, els paràmetres cinemàtics (CAD, OV, TCT i LG) i l’SmO2.
Es va fer un ANOVA unifactorial independent per examinar les diferències als índexs d’asimetria de la PD, els paràmetres cinemàtics (CAD, OV, TCT i LG) i l’SmO2 entre nivells d’intensitat. L’asimetria (%) de cada variable es va calcular d’acord amb la següent equació:
on Xr representa l’SmO2 registrada en la cama dreta i Xl representa l’SmO2 corresponent registrada en la cama esquerra. Aquest enfocament segueix protocols establerts (Karamanidis et al., 2003; Knapik et al., 1991).
Tant en l’ANOVA de mesures repetides de dos factors com en l’ANOVA unifactorial dels índexs d’asimetria, es van fer comparacions per parells post hoc utilitzant el mètode de correcció d’Holm per controlar la taxa d’error familiar deguda a les comparacions múltiples, i es notifiquen els valors de p ajustats. Per a les comparacions per parells posteriors a l’ANOVA de dos factors, les mides de l’efecte es van calcular mitjançant la d de Cohen, amb valors de 0.2, 0.5 i 0.8 interpretats com a efectes petits, mitjans i grans, respectivament (Cohen, 1988). Per a l’ANOVA unifactorial aplicat als índexs d’asimetria amb distribució normal (p. ex., SmO₂ ASIM), les mides de l’efecte es van estimar mitjançant eta al quadrat parcial (η²p), amb llindars de .01, .06 i .14 considerats efectes petits, mitjans i grans, respectivament (Richardson, 2011). En canvi, per als índexs d’asimetria que no van complir el supòsit de normalitat (p. ex., PD ASIM, CAD ASIM, OV ASIM, TCT ASIM i LG ASIM), es va aplicar la prova de Friedman, seguida de proves de rangs amb signe de Wilcoxon i correcció d’Holm per a les comparacions per parells; en aquests casos, les mides de l’efecte es van calcular mitjançant la W de Kendall, interpretada com a nivells de concordança petits (.1), moderats (.3) i grans (.5) (Tomczak i Tomczak, 2014).
Totes les anàlisis estadístiques es van fer amb Microsoft Excel (versió 16.81, 24011420) i JASP (versió 0.18.3). La significació estadística es va fixar en p < .05 per als efectes globals.
Resultats
Els participants (n = 14) van assolir una freqüència cardíaca màxima (FCmáx) de 183.5 ± 15.6 lpm i una VAM de 18.53 ± 1.02 km·h-1 durant el test de VAM-EVAL. L’SmO2, la PD i els paràmetres cinemàtics van mostrar canvis progressius en resposta a l’augment de la velocitat de cursa al llarg de la prova.
Durant les etapes incrementals del test de VAM-EVAL (60 %, 70 %, 80 % i 90 % de la VAM), la PD, el CAD i l’LG van augmentar significativament a ambdues cames en totes les intensitats, mentre que el TCT va disminuir significativament en totes les intensitats (totes les p. inferiors a .05) (Taula 2). Es va observar un efecte principal significatiu de la variable Cama en la PD (F(1,13) = 13.62, p = .003, η²p = .512), l’OV (F(1,13) = 7.97, p = .014, η²p = .38), el TCT (F(1,13) = 6.51, p = .024, η²p = .334) i l’LG (F(1,13) = 21.26, p < .001, η²p = .621). Tanmateix, les comparacions post hoc entre la cama interior i l’exterior en cada nivell d’intensitat només van assolir significació estadística després de la correcció d’Holm en la PD al 80 % de la VAM (t(13) = −4.455, p = .003, d = −0.29), l’LG al 70 % de la VAM (t(13) = −6.298, p < .001, d = -0.37)i l’LG al 80 % de la VAM (t(13) = −4.717, p < .001, d = -0.47). Això suggereix un efecte principal constant de la variable Cama en l’OV i el TCT, amb diferències sistemàtiques d’una cama respecte de l’altra en totes les intensitats. Tanmateix, l’absència d’una interacció significativa entre Cama i Intensitat indica que la magnitud d’aquesta diferència entre extremitats es va mantenir estable, independentment del nivell d’intensitat. No es van trobar diferències significatives entre cames en el CAD (p = .313, η²p = .08) ni en l’SmO₂ (p = .976, η²p = .00).
Taula 2
Dades descriptives d’ambdues cames en PD, CAD, OV, TCT, LG i SmO₂ durant el test de VAM-EVAL (n = 14; s’indiquen les diferències significatives (p < .05) entre nivells d’intensitat)
L’anàlisi de la mida de l’efecte va revelar diferències entre extremitats entre moderades i grans en la PD, l’OV, el TCT i l’LG (η²p entre .33 i .62). En canvi, el TCT, l’LG i la PD van mostrar adaptacions intramembre marcades amb l’augment de la intensitat, tal com reflecteixen els significatius efectes d’intensitat i els valors d’eta al quadrat parcial superiors a .95 (Taula 3). Tant la cama interior com l’exterior van mostrar mides de l’efecte intramembre molt grans en resposta a l’augment de la intensitat, especialment en el TCT, l’LG i la PD. Les mides de l’efecte van oscil·lar entre 1.07 i 3.26 en la PD, entre 1.75 i 6.97 en el TCT, i entre 1.38 i 4.76 en l’LG. El CAD i l’SmO₂ també van mostrar canvis intramembre, encara que de menor magnitud. Les mides de l’efecte van oscil·lar entre 0.52 i 2.16 en el CAD i entre 0.08 i 1.15 en l’SmO₂, la qual cosa suggereix ajustaments intramembre entre moderats i grans, especialment en els nivells d’intensitat més alts. En canvi, l’OV només va mostrar mides de l’efecte petites i inconsistents (d = 0.08 a 0.39), la qual cosa indica una resposta limitada a l’augment de la intensitat.
Taula 3
Resultats de l’ANOVA de mesures repetides de dos factors per a PD, CAD, OV, TCT, LG i SmO₂ durant el test de VAM-EVAL (n = 14)
Quant als valors d’asimetria entre extremitats, no es van observar diferències significatives en la PD, el CAD, l’LG, l’OV ni el TCT entre els diferents nivells d’intensitat (Taula 4). Tanmateix, l’asimetria en l’SmO₂ va mostrar diferències significatives entre el 60 % i el 90 % i entre el 70 % i el 90 % de la VAM (p < .05).
Taula 4
Dades descriptives dels índexs d’asimetria de PD, CAD, OV, TCT, LG i SmO2 entre nivells d’intensitat (60 %, 70 %, 80 % i 90 % de la VAM; s’indiquen les diferències significatives entre nivells d’intensitat [p < .05])
L’anàlisi de la mida de l’efecte va revelar efectes entre petits i grans en l’asimetria de l’SmO₂ entre intensitats, que van oscil·lar entre petits (60-70 % de la VAM: d = 0.081; 70-80 % de la VAM: d = 0.266), moderats (60-80 % de la VAM: d = 0.347; 80-90 % de la VAM: d = 0.574) i grans (60-90 % de la VAM: d = 0.921; 70-90 % de la VAM: d = 0.840) (Taula 4). Aquests resultats suggereixen que l’asimetria d’SmO₂ augmenta progressivament a mesura que s’incrementa la intensitat de cursa.
Taula 5
Resultats de les anàlisis estadístiques dels índexs d’asimetria entre extremitats entre nivells d’intensitat durant el test de VAM-EVAL
La Figura 1 il·lustra els patrons d’SmO₂ d’ambdues cames en dos participants representatius. El plafó esquerre mostra un participant amb asimetries marcades, mentre que el dret en mostra un sense asimetria observable. En el participant asimètric, encara que els valors absoluts d’SmO₂ difereixen considerablement entre cames, els perfils temporals de saturació continuen sent notablement similars entre intensitats.
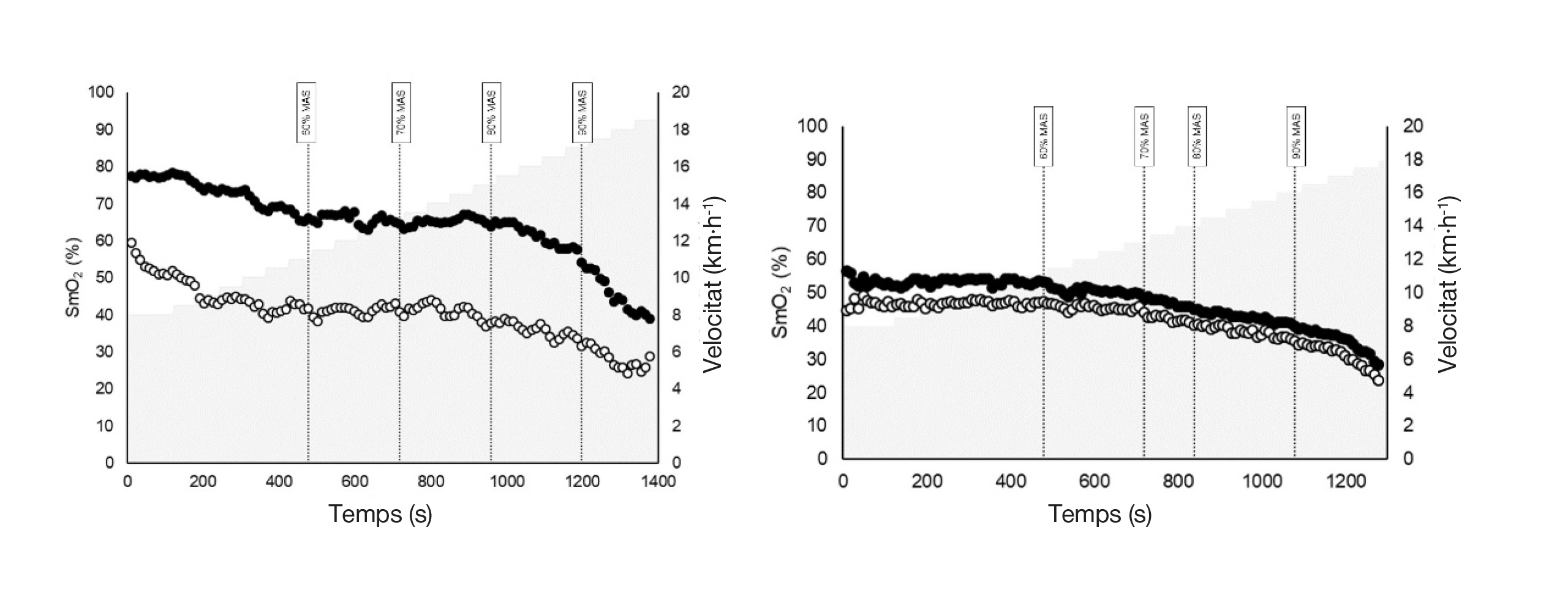
Nota. Els valors de SmO₂ s’expressen en forma de percentatge (%), i la velocitat de cursa s’expressa en quilòmetres per hora (km·h-1). La línia negra representa la cama exterior, mentre que la línia de color clar representa la cama interior. VAM = velocitat aeròbica màxima.
Discussió
El present estudi és un dels primers en analitzar les asimetries de potència, cinemàtica i oxigenació muscular entre la cama interior i l’exterior durant una cursa incremental en pista de 400 m en un entorn realista a l’aire lliure. La majoria dels estudis previs d’aquest àmbit s’han fet en condicions controlades de laboratori en interiors, que potser no captin plenament la complexitat biomecànica i fisiològica de la cursa en condicions reals.
El primer objectiu era determinar si córrer sobre una pista de 400 m indueix diferències mesurables en la PD, les variables cinemàtiques (CAD, OV, TCT, LG) i l’SmO₂ entre la cama interior i l’exterior. Les principals observacions van revelar efectes principals significatius de la variable Cama en la PD, l’LP, l’OV i el TCT, fet que indica diferències entre extremitats sistemàtiques en tots els nivells d’intensitat. Tanmateix, les anàlisis post hoc només van identificar diferències amb significació estadística en determinades intensitats (PD al 80 % de la VAM, i LG al 70 % i al 80 % de la VAM), mentre que no es van observar diferències entre extremitats significatives en el CAD ni en l’SmO₂. A més, l’absència d’interaccions significatives entre Cama i Intensitat suggereix que aquestes asimetries es van mantenir estables al llarg del protocol incremental, en comptes d’augmentar amb la intensitat de l’exercici.
El segon objectiu va ser avaluar la magnitud i la progressió de les asimetries entre extremitats a mesura que augmentava la intensitat de cursa. Les principals troballes van mostrar que els índexs d’asimetria de la PD, el CAD, l’OV, el TCT i l’LG no van canviar significativament entre els nivells d’intensitat, la qual cosa suggereix una simetria mecànica estable al llarg de la prova. En canvi, l’asimetria d’SmO₂ va augmentar progressivament amb la intensitat, amb mides de l’efecte grans observades al 90 % de la VAM. Aquesta divergència fisiològica suggereix un desequilibri creixent en l’oxigenació muscular local entre extremitats sota un estrès metabòlic més gran; això posiciona l’SmO₂ com un marcador sensible i potencialment precoç d’asimetria dependent de la càrrega durant la cursa a alta velocitat.
Aquestes observacions coincideixen parcialment amb estudis previs que van notificar alteracions cinemàtiques durant la cursa en corba; tanmateix, a l’hora de comparar els resultats s’han de tenir en compte algunes diferències metodològiques importants. Per exemple, els estudis de Chang i Kram (2007) i Hamill et al. (1987) van demostrar que la cursa en corba afecta variables cinemàtiques rellevants com el CAD, l’LG, l’FP, l’OV i el TCT. És important reconèixer les diferències considerables entre les seves condicions experimentals i les del present estudi. En concret, Chang i Kram (2007) van investigar la cursa en corbes amb un radi molt menor (6 m) que el d’una pista d’atletisme estàndard (36.5 m), i els seus participants van córrer a velocitats màximes d’esprint, a diferència de les intensitats submàximes utilitzades en el nostre protocol (fins al 100 % de la VAM; 5.15 ± 0.28 m·s-1). De la mateixa manera, Hamill et al. (1987) van examinar la cursa a una velocitat considerablement superior (6.31 ± 5 m·s-1) al ritme mantingut pels triatletes en el nostre estudi. Aquestes diferències metodològiques van produir forces centrífugues molt més grans, la qual cosa va amplificar les disparitats cinemàtiques que van constatar entre la cama interior i l’exterior.
També observem que, a mesura que augmentava la intensitat de la cursa, es produïen increments significatius en la PD, el CAD i l’LG en ambdues cames en tots els nivells d’intensitat. En concret, la PD en la CI va augmentar un 13.6 %, un 13.2 % i un 10.4 % entre el 60 % i el 70 %, entre el 70 % i el 80 %, i entre el 80 % i el 90 % de la VAM, respectivament. De forma similar, la PD en la CE va augmentar un 13.9 %, un 11.2 % i un 11.9 % en els mateixos intervals. Quant al CAD, les dues cames van mostrar increments moderats però constants amb la intensitat. El CAD en la CI va augmentar un 2.1 %, un 2.2 % i un 3.4 %, mentre que la CE va mostrar increments lleugerament menors però progressius del 1.7 %, el 2.4 % i el 3.6 % en els intervals corresponents. L’LG també va augmentar amb la intensitat, encara que la taxa d’increment va disminuir en apropar-se al 90 % de la VAM. En la CI, l’LG va augmentar un 13.7 %, un 11.5 % i un 8.0 %, mentre que en la CE es van observar increments del 13.2 %, l’11.1 % i el 9.3 %. Al contrari, el TCT va disminuir de forma constant en les dues cames a mesura que va augmentar la intensitat. En la CI, el TCT es va reduir un 9.8 %, un 8.6 % i un 6.8 % entre el 60 % i el 70 %, entre el 70 % i el 80 %, i entre el 80 % i el 90 % de la VAM, respectivament. En la CE, les reduccions van ser del 9.8 %, el 8.9 % i el 7.6 % en els mateixos intervals d’intensitat. Aquestes observacions coincideixen amb les de Patoz et al. (2023), especialment quant a la coherència de les respostes biomecàniques a mesura que augmenten les intensitats de cursa. Igual que en les seves observacions al 90 %-120 % de la velocitat aeròbica de pic, les nostres dades no van revelar interaccions significatives entre Cama i Intensitat (p > .05), fet que indica un patró d’adaptació uniforme en ambdues extremitats. A més, ambdós estudis van notificar una cadència (freqüència de gambada) i una oscil·lació vertical estables malgrat l’augment de la fatiga, cosa que suggereix que els corredors prioritzen el manteniment del ritme de moviment i el control del centre de masses. L’efecte significatiu de la intensitat observat en les nostres dades de temps de contacte amb el terra també reforça la seva interpretació que els corredors adopten estratègies espaciotemporals, com la modulació del temps de contacte, per optimitzar la distribució de la càrrega durant esforços d’alta intensitat. Si bé ambdós estudis es van centrar en les respostes biomecàniques a intensitats submàximes i properes a la màxima, el nostre protocol va ser lleugerament menys exigent. En el nostre estudi, els esportistes van fer una prova incremental contínua sense períodes de descans, mentre que Patoz et al. (2023) van dur a terme quatre assajos exhaustius independents a percentatges fixos de la velocitat de pic de cada participant, efectuats en ordre aleatoritzat.
A més dels ajustaments mecànics observats en els paràmetres espaciotemporals, la resposta fisiològica muscular també va mostrar tendències rellevants a mesura que va augmentar la intensitat. En concret, la saturació muscular d’oxigen (SmO₂) va disminuir progressivament tant en la cama interior com en l’exterior al llarg del protocol incremental. Aquesta resposta encaixa amb estudis previs sobre oxigenació muscular durant exercici graduat, que van notificar una disminució contínua de l’SmO₂ a causa de l’augment de la demanda d’extracció d’oxigen (Grassi et al., 1999). Les presents observacions confirmen aquest patró, especialment a intensitats més altes, com el 90 % de la VAM.
No es van observar diferències significatives entre la cama interior i l’exterior (p = .976), ni hi va haver una interacció significativa entre Cama i Intensitat (p = .068). Tanmateix, es va observar un fort efecte principal de la intensitat (p < .001, η²p = .697), fet que indica que l’SmO₂ va respondre de forma marcada als augments de la velocitat de cursa. Aquests resultats suggereixen que els canvis observats en l’SmO₂ reflecteixen una adaptació fisiològica global a l’augment de la demanda metabòlica, sense una evidència clara de desequilibris funcionals entre la cama interior i l’exterior quant a l’oxigenació muscular.
L’objectiu secundari d’aquest estudi era avaluar la magnitud de les asimetries de la PD, les variables cinemàtiques (CAD, OV, TCT i LG) i l’SmO2 en triatletes de nivell nacional durant un test de VAM-EVAL. Les principals observacions no van mostrar diferències significatives entre intensitats, excepte en l’SmO2, que va presentar canvis significatius entre el 60 % i el 90 % i entre el 70 % i el 90 % de la VAM. Aquests resultats suggereixen que l’asimetria d’SmO₂ és proporcional a la intensitat de l’exercici, especialment al 90 % de la VAM, la qual cosa indica que, a mesura que augmenta la intensitat de cursa, les diferències en la utilització d’oxigen entre una cama i l’altra es fan més evidents en els triatletes.
Per contextualitzar millor aquestes conclusions, és essencial considerar els mecanismes fisiològics subjacents que podrien explicar les asimetries observades. La presència d’una asimetria significativa d’SmO₂, malgrat una producció mecànica simètrica en paràmetres com la potència i la longitud de gambada, apunta al paper de factors musculars intrínsecs. En particular, les variacions locals en la capil·larització i les adaptacions vasculars podrien contribuir a aquestes disparitats. La capil·larització és un factor determinant de l’aportació i la difusió d’oxigen; concretament, paràmetres com la densitat capil·lar i el nombre de contactes capil·lars per àrea de fibra (CC/AF) s’associen amb una extracció d’O₂ més eficient; aquesta relació és especialment sòlida entre els joves (Chilibeck et al., 1997). Així mateix, les adaptacions vasculars provocades per l’entrenament de resistència solen ser localitzades, amb una resposta microvascular més gran principalment en els teixits directament implicats en l’esforç (Soares et al., 2018).
Els processos fisiològics interns poden continuar sent asimètrics a causa de diferències en la perfusió muscular local o el metabolisme oxidatiu. En aquest context, determinats mecanismes reflexos del sistema nerviós autònom, com el reflex pressor de l’exercici, poden augmentar la irrigació sanguínia dels músculs que fan un esforç més gran, la qual cosa podria reduir la disponibilitat d’oxigen a altres músculs menys implicats en la tasca o amb una funció auxiliar. Aquests mecanismes formen part de la resposta autònoma general a l’augment de la demanda metabòlica i participen en la modulació de les respostes vasculars específiques de cada extremitat durant l’exercici intens (Orcioli-Silva et al., 2024).
A més, estratègies neuromusculars com la coactivació muscular contribueixen a aquestes asimetries internes. A velocitats de cursa elevades, la coactivació global augmenta i es produeix abans i durant períodes més breus per incrementar la rigidesa de tota l’extremitat i l’estabilització articular (Fiori et al., 2024). Es pot destacar que la relació entre excitació muscular (RMS) i SmO₂ no és uniforme a tots els músculs. Mentre que els músculs generadors de potència, com el vast lateral, mostren un patró invers clar (l’RMS augmenta mentre l’SmO₂ disminueix) a causa d’una demanda metabòlica més gran, alguns estabilitzadors, com el gastrocnemi medial, mantenen un nivell estable d’SmO₂ i d’RMS durant tot l’exercici (Sendra-Pérez et al. 2025). Això reflecteix les seves funcions diferenciades, ja que els músculs estabilitzadors participen més en la regulació de la càrrega que en la generació de força. Aquestes observacions posen en relleu la complexa interacció entre mecanismes neuromusculars i vasculars que modulen la dinàmica de l’SmO₂. Atès que el present estudi es va centrar exclusivament en el vast lateral, l’anàlisi d’altres músculs amb funcions diverses permetria conèixer de forma més completa aquestes asimetries fisiològiques.
Malgrat que la majoria dels estudis anteriors ha avaluat la simetria d’SmO₂ mitjançant mètodes estadístics de concordança i diferències entre extremitats absolutes, la nostra anàlisi va fer servir un índex d’asimetria normalitzat (ASI%) per expressar la disparitat relativa entre cames. En condicions controlades de laboratori, Sendra-Pérez et al.(2025) van definir la simetria a partir de diferències absolutes d’SmO₂ d’entre el 10 % i el 20 %, mentre que Skotzke et al. (2024) van utilitzar l’anàlisi de Bland-Altman per establir límits de concordança de ± 20 %, i van concloure que aquestes diferències entre extremitats són freqüents i no estan vinculades a la dominància de la cama. En canvi, els nostres valors d’ASI% durant la cursa a l’aire lliure van oscil·lar entre el 24.4 % i el 53.5 %, per sobre tant dels llindars de laboratori com del canvi mínim detectable (CMD) del 18 % notificat per Skotzke et al. (2024) per a l’SmO₂ en el ciclisme.
Tanmateix, és important assenyalar que l’ASI% es veu influït pels valors absoluts baixos. Atès que els nivells d’SmO₂ van descendir de forma marcada a intensitats elevades (p. ex., ~25 % al 90 % de la VAM), fins i tot diferències absolutes moderades poden generar valors d’ASI% desproporcionadament alts. Per validar les nostres observacions, vam fer una anàlisi de Bland-Altman en totes les intensitats. Els resultats van revelar un biaix sistemàtic mínim al 60 %, 70 % i 80 % de la VAM (−2.6 % ± 24.0 %, +0.2 % ± 24.3 % i +0.3 % ± 28.3 %, respectivament), amb límits de concordança relativament coherents. Al 90 % de la VAM, malgrat que els valors d’SmO₂ van ser més baixos, el biaix va continuar sent petit (+1.7 %) i els límits de concordança (± 25.68 %) no es van ampliar de forma significativa en comparació amb les etapes anteriors. En comparació amb Skotzke et al. (2024), que van notificar límits de concordança de ± 20 % durant l’esforç estable amb bicicleta, els nostres resultats mostren nivells d’asimetria fisiològica lleugerament més amplis, però encara comparables en condicions de cursa sobre el terreny més exigents. Aquests resultats suggereixen que, si bé les asimetries fisiològiques d’SmO₂, tal com les capta l’ASI%, sí que es fan més evidents amb l’augment de la intensitat, la magnitud de l’asimetria relativa s’ha d’interpretar amb cautela, especialment en condicions de baixa saturació d’oxigen.
Segons la recent revisió sistemàtica de D’Hondt et al. (2024), la relació entre l’asimetria entre extremitats i el rendiment en cursa de resistència és complexa. Les seves observacions indiquen que alguns indicadors d’asimetria funcionals, morfològics, cinemàtics i cinètics, encara que no tots, són inversament proporcionals al rendiment en cursa o no hi tenen una relació significativa. En el present estudi, malgrat mostrar asimetries significatives d’SmO₂, especialment a intensitats elevades, els triatletes van assolir un nivell de rendiment de 3, segons la classificació proposada per De Pauw et al. (2013). Per analitzar les asimetries en més profunditat, hauria estat beneficiós fer una avaluació preliminar de la composició corporal mitjançant absorciometria de raigs X de doble energia (DXA) per avaluar la composició corporal total i identificar possibles diferències de massa muscular entre cames. Branski et al. (2010) van demostrar que l’asimetria de massa magra influeix en l’asimetria de força i potència durant tasques de salt en esportistes universitaris. Així mateix, la realització de proves específiques de força unilateral per a corredors podria haver proporcionat informació sobre possibles desequilibris de força entre extremitats. Aquest plantejament hauria permès una interpretació més completa de les diferències observades en la saturació muscular d’oxigen.
Limitacions i futurs estudis
Aquest estudi presenta algunes limitacions que s’han de tenir en compte. En primer lloc, la mida de la mostra va ser relativament reduïda i va incloure únicament triatletes homes de nivell nacional, la qual cosa podria limitar la generalització de les observacions a altres poblacions, com són corredores professionals o persones que practiquen aquest esport de manera informal. En segon lloc, l’SmO2 es va mesurar exclusivament en el vast lateral. La inclusió d’altres músculs implicats en la cursa podria haver permès caracteritzar d’una forma més completa les asimetries fisiològiques entre extremitats. En tercer lloc, no es van incloure mesures directes de força muscular unilateral, funció neuromuscular o asimetria de la composició corporal, la qual cosa podria haver contribuït a una interpretació més profunda de les asimetries d’SmO₂ observades. Finalment, encara que el fet de fer la prova a l’aire lliure apropa l’avaluació a les condicions reals de cursa, també podria haver introduït més variabilitat que un entorn de laboratori.
Els futurs estudis haurien de continuar posant èmfasi en entorns realistes per reflectir amb més precisió les demandes de la cursa a l’aire lliure. Així mateix, la integració d’anàlisis de composició corporal i avaluacions específiques de força d’ambdues cames podria ajudar a identificar els orígens fisiològics de les asimetries d’SmO₂. L’ampliació d’aquests estudis contribuiria a conèixer millor els mecanismes fisiològics i biomecànics subjacents als desequilibris entre extremitats i les seves possibles conseqüències per al rendiment esportiu. Finalment, els futurs estudis haurien d’incorporar eines d’observació sistemàtica i anàlisis amb vídeo (Soto et al., 2019) per augmentar la fiabilitat de l’anàlisi del moviment en entorns esportius aplicats.
Conclusions
Les conclusions d’aquest estudi en triatletes homes de nivell nacional que van fer una prova incremental de cursa sobre pista de 400 m indiquen que, encara que no es van observar asimetries en la potència desenvolupada ni en els paràmetres cinemàtics, les asimetries de saturació muscular d’oxigen es van fer evidents a mida que va augmentar la intensitat de cursa. Aquestes asimetries no van ser atribuïbles a la trajectòria de cursa en corba, ja que no es van trobar diferències significatives entre la cama interior i l’exterior, excepte en la potència de sortida al 80 % de la velocitat aeròbica màxima (VAM) i la longitud de gambada al 70 % i al 80 % de la VAM. Si bé els paràmetres mecànics es van mantenir en gran manera simètrics, la divergència progressiva en els valors d’SmO₂ apunta a un desequilibri fisiològic que s’accentua a intensitats més altes, la qual cosa destaca l’SmO₂ com a marcador sensible d’asimetries internes dependents de la càrrega, fins i tot en absència de discrepàncies mecàniques.
Agraïments
Els autors desitgen agrair a l’Institut Nacional d’Educació Física de Catalunya i al Grup d’Investigació en Ciències de l’Esport INEFC Barcelona (GRCEIB, https://inefc-grceib.cat) el seu suport institucional, així com a Josep Tarrés la seva col·laboració en el procés de recollida de dades.
Els autors declaren que no es van utilitzar eines d’intel·ligència artificial generativa a la redacció, l’anàlisi de les dades ni la interpretació d’aquest manuscrit, d’acord amb les polítiques de publicació d’Apunts.
Finançament
Aquest estudi va ser finançat per l’Institut Nacional d’Educació Física de Catalunya (INEFC) de la Generalitat de Catalunya. L’entitat finançadora no va participar en el disseny de l’estudi ni en la recollida i l’anàlisi de les dades ni en la decisió de publicar ni en la preparació del manuscrit.
Referències
[1] Alt, T., Heinrich, K., Funken, J., & Potthast, W. (2015). Lower extremity kinematics of athletics curve sprinting. Journal of Sports Sciences, 33(6), 552–560. doi.org/10.1080/02640414.2014.960881
[2] Bell, D. R., Sanfilippo, J. L., Binkley, N., & Heiderscheit, B. C. (2014). Lean mass asymmetry influences force and power asymmetry during jumping in collegiate athletes. Journal of Strength and Conditioning Research, 28(4), 884–891. doi.org/10.1519/JSC.0000000000000367
[3] Bini, R., & Hume, P. (2015). Relationship between pedal force asymmetry and performance in cycling time trial. Journal of Sports Medicine and Physical Fitness, 55(9), 892–898.
[4] Bishop, C., Read, P., Brazier, J., Jarvis, P., Chavda, S., Bromley, T., & Turner, A. (2021). Effects of interlimb asymmetries on acceleration and change of direction speed: A between-sport comparison of professional soccer and cricket athletes. Journal of Strength and Conditioning Research, 35(8), 2095–2101. doi.org/10.1519/JSC.0000000000003135
[5] Bishop, C., Read, P., Bromley, T., Brazier, J., Jarvis, P., Chavda , S., & Turner, A. (2022). The association between interlimb asymmetry and athletic performance tasks: A season-long study in elite academy soccer players. Journal of Strength and Conditioning Research, 36(3), 787–795. doi.org/10.1519/JSC.0000000000003526
[6] Bishop, C., Turner, A., & Read, P. (2017). Effects of inter-limb asymmetries on physical and sports performance: A systematic review. Journal of Sports Sciences, 36(10), 1135–1144. doi.org/10.1080/02640414.2017.1361894
[7] Branski, L. K., Norbury, W. B., Herndon, D. N., Chinkes, D. L., Cochran, A., Suman, O., Benjamin, D., & Jeschke, M. G. (2010). Measurement of body composition in burned children: Is there a gold standard? JPEN: Journal of Parenteral and Enteral Nutrition, 34(1), 55–63. doi.org/10.1177/0148607109336601
[8] Cerezuela-Espejo, V., Hernández-Belmonte, A., Courel-Ibáñez, J., Conesa-Ros, E., Mora-Rodríguez, R., & Pallarés, J. G. (2021). Are we ready to measure running power? Repeatability and concurrent validity of five commercial technologies. European Journal of Sport Science, 21(3), 341–350. doi.org/10.1080/17461391.2020.1748117
[9] Chang, Y. H., & Kram, R. (2007). Limitations to maximum running speed on flat curves. Journal of Experimental Biology, 210(6), 971–982. doi.org/10.1242/jeb.02728
[10] Chilibeck, P. D., Paterson, D. H., Cunningham, D. A., Taylor, A. W., & Noble, E. G. (1997). Muscle capillarization, O2 diffusion distance, and VO₂ kinetics in old and young individuals. Journal of Applied Physiology, 82(1), 63–69. doi.org/10.1152/jappl.1997.82.1.63
[11] Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). Routledge. doi.org/10.4324/9780203771587
[12] Connick, M. J., & Li, F. X. (2015). Prolonged cycling alters stride time variability and kinematics of a post-cycle transition run in triathletes. Journal of Electromyography and Kinesiology, 25(1), 34–39. doi.org/10.1016/j.jelekin.2014.08.009
[13] D’Hondt, J., Chapelle, L., Bishop, C., Aerenhouts, D., Pauw, K.D., Clarys, P., & D’Hondt, E. (2024). Association between inter-limb asymmetry and endurance running performance in healthy populations: A systematic review. Sports Med - Open, 10(127). doi.org/10.1186/s40798-024-00790-w
[14] Feldmann, A., Schmitz, R., & Erlacher, D. (2019). Near-infrared spectroscopy-derived muscle oxygen saturation on a 0% to 100% scale: Reliability and validity of the Moxy Monitor. Journal of Biomedical Optics, 24(11), 115001. doi.org/10.1117/1.JBO.24.11.115001
[15] Fiori, L., Castiglia, S. F., Chini, G., Draicchio, F., Sacco, F., Serrao, M., Tatarelli, A., Varrecchia, T., & Ranavolo, A. (2024). The lower limb muscle co-activation map during human locomotion: From slow walking to running. Bioengineering, 11(3), 288. doi.org/10.3390/bioengineering11030288
[16] Fox, K. T., Pearson, L. T., & Hicks, K. M. (2023). The effect of lower inter-limb asymmetries on athletic performance: A systematic review and meta-analysis. PLoS One, 18(6), e0286942. doi.org/10.1371/journal.pone.0286942
[17] García, G. C., & Secchi, J. D. (2013). Relationship between the final speeds reached in the 20 metre Course Navette and the MAS-EVAL test. A proposal to predict the maximal aerobic speed. Apunts Medicina de l’Esport, 48(177), 27–34. doi.org/10.1016/j.apunts.2011.11.004
[18] Gilgen-Ammann, R., Taube, W., & Wyss, T. (2017). Gait asymmetry during 400- to 1000-m high-intensity track running in relation to injury history. International Journal of Sports Physiology and Performance, 12(S2), S2-157-S2-160. doi.org/10.1123/ijspp.2016-0379
[19] Grassi, B., Quaresima, V., Marconi, C., Ferrari, M., & Cerretelli, P. (1999). Blood lactate accumulation and muscle deoxygenation during incremental exercise. Journal of Applied Physiology, 87(1), 348–355. doi.org/10.1152/jappl.1999.87.1.348
[20] Hamill, J., Murphy, M., & Sussman, D. (1987). The effects of track turns on lower extremity function. International Journal of Sport Biomechanics, 3(3), 276–286. doi.org/10.1123/ijsb.3.3.276
[21] Heiden, T., & Burnett, A. (2003). The effect of cycling on muscle activation in the running leg of an Olympic distance triathlon. Sports Biomechanics, 2(1), 35–49. doi.org/10.1080/14763140308522806
[22] Helme, M., Tee, J., Emmonds, S., & Low, C. (2021). Does lower-limb asymmetry increase injury risk in sport? A systematic review. Physical Therapy in Sport, 49, 204–213. doi.org/10.1016/j.ptsp.2021.03.001
[23] Jacques, T., Bini, R., & Arndt, A. (2021). Running after cycling induces inter-limb differences in muscle activation but not in kinetics or kinematics. Journal of Sports Sciences, 39(2), 154–160. doi.org/10.1080/02640414.2020.1809176
[24] Karamanidis, K., Arampatzis, A., & Brüggemann, G.-P. (2003). Symmetry and reproducibility of kinematic parameters during various running techniques. Medicine and Science in Sports and Exercise, 35(6), 1009–1016. doi.org/10.1249/01.MSS.0000069337.49567.F0
[25] Knapik, J. J., Bauman, C. L., Jones, B. H., Harris, J. M., & Vaughan, L. (1991). Preseason strength and flexibility imbalances associated with athletic injuries in female collegiate athletes. American Journal of Sports Medicine, 19(1), 76–81. doi.org/10.1177/036354659101900113
[26] Léger, L., & Boucher, R. (1980). An indirect continuous running multistage field test: the Université de Montréal track test. Canadian journal of applied sport sciences. Journal canadien des sciences appliquées au sport, 5(2), 77–84.
[27] Loturco, I., Pereira, L. A., Kobal, R., Abad, C. C. C., Rosseti, M., Carpes, F. P., & Bishop, C. (2019). Do asymmetry scores influence speed and power performance in elite female soccer players? Biology of Sport, 36(3), 209–216. doi.org/10.5114/biolsport.2019.85454
[28] McManus, C. J., Collison, J., & Cooper, C. E. (2018). Performance comparison of the MOXY and PortaMon near-infrared spectroscopy muscle oximeters at rest and during exercise. Journal of Biomedical Optics, 23(1), 015007. doi.org/10.1117/1.JBO.23.1.015007
[29] Millet, G. P., & Vleck, V. E. (2000). Physiological and biomechanical adaptations to the cycle to run transition in Olympic triathlon: review and practical recommendations for training. British journal of sports medicine, 34(5), 384–390. doi.org/10.1136/bjsm.34.5.384
[30] Olcina, G., Perez-Sousa, M. Á., Escobar-Alvarez, J. A., & Timón, R. (2019). Effects of cycling on subsequent running performance, stride length, and muscle oxygen saturation in triathletes. Sports, 7(5), 115. doi.org/10.3390/sports7050115
[31] Orcioli-Silva, D., Beretta, V. S., Santos, P. C. R., Rasteiro, F. M., Marostegan, A. B., Vitório, R., Gobatto, C. A., & Manchado-Gobatto, F. B. (2024). Cerebral and muscle tissue oxygenation during exercise in healthy adults: A systematic review. Journal of sport and health science, 13(4), 459–471. doi.org/10.1016/j.jshs.2024.03.003
[32] Patoz, A., Blokker, T., Pedrani, N., Spicher, R., Borrani, F., & Malatesta, D. (2023). Biomechanical adaptations during exhaustive runs at 90 to 120% of peak aerobic speed. Scientific Reports, 13, 8236. doi.org/10.1038/s41598-023-35345-8
[33] Pauw, K.D., Roelands, B., Cheung, S. S., de Geus, B., Rietjens, G., & Meeusen, R. (2013). Guidelines to classify subject groups in sport-science research. International Journal of Sports Physiology and Performance, 8(2), 111–122. doi.org/10.1123/ijspp.8.2.111
[34] Richardson, J. T. (2011). Eta squared and partial eta squared as measures of effect size in educational research. Educational research review, 6(2), 135–147. doi.org/10.1016/j.edurev.2010.12.001
[35] Sendra-Pérez, C., Priego-Quesada, J. I., Murias, J. M., Carpes, F. P., Salvador-Palmer, R., & Encarnación-Martínez, A. (2025). Evaluation of leg symmetry in muscle oxygen saturation during submaximal to maximal cycling exercise. European Journal of Sport Science, 25(1), e12230. doi.org/10.1002/ejsc.12230
[36] Skotzke, P., Schwindling, S., & Meyer, T. (2024). Side differences and reproducibility of the Moxy muscle oximeter during cycling in trained men. European Journal of Applied Physiology, 124, 3075–3083. doi.org/10.1007/s00421-024-05514-2
[37] Soares, R. N., George, M. A., Proctor, D. N., & Murias, J. M. (2018). Differences in vascular function between trained and untrained limbs assessed by near-infrared spectroscopy. European Journal of Applied Physiology, 118, 2241–2248. doi.org/10.1007/s00421-018-3955-3
[38] Soto, A., Camerino, O., Iglesias, X., Anguera, M. T., & Castañer, M. (2019). LINCE PLUS: Research software for behaviour video analysis. Apunts. Educación Física y Deportes, 137, 149–153. doi.org/10.5672/apunts.2014-0983.es.(2019/3).137.11
[39] Tomczak, E. & Tomczak, M. (2014). The need to report effect size estimates revisited. An overview of some recommended measures of effect size. TRENDS in Sport Sciences, 21(1).
[40] van der Zwaard, S., Jaspers, R. T., Blokland, I. J., Achterberg, C., Visser, J. M., den Uil, A. R., Hofmijster, M. J., Levels, K., Noordhof, D. A., de Haan, A., de Koning, J. J., van der Laarse, W. J., & de Ruiter, C. J. (2016). Oxygenation Threshold Derived from Near-Infrared Spectroscopy: Reliability and Its Relationship with the First Ventilatory Threshold. PloS one, 11(9), e0162914. doi.org/10.1371/journal.pone.0162914
[41] van Rassel, C. R., Ajayi, O. O., Sales, K. M., Griffiths, J. K., Fletcher, J. R., Edwards, W. B., & MacInnis, M. J. (2023). Is running power a useful metric? Quantifying training intensity and aerobic fitness using Stryd running power near the maximal lactate steady state. Sensors, 23(21), 8729. doi.org/10.3390/s23218729
[42] Vasquez-Bonilla, A. A., Brazo-Sayavera, J., Timón, R., & Olcina, G. (2022). Monitoring muscle oxygen asymmetry as a strategy to prevent injuries in footballers. Research Quarterly for Exercise and Sport, 94(3), 609–617. doi.org/10.1080/02701367.2022.2026865
[43] Vasquez-Bonilla, A., Tomas-Carus, P., Brazo-Sayavera, J., Malta, J., Folgado, H., & Olcina, G. (2023). Muscle oxygenation is associated with bilateral strength asymmetry during isokinetic testing in sport teams. Science & Sports, 38(4), 426.e1–426.e9. doi.org/10.1016/j.scispo.2022.03.014
[44] World Athletics. (2019). Track and field facilities manual: Chapters 1–3. World Athletics. worldathletics.org/about-iaaf/documents/technical-information
[45] Yanci, J. (2014). Muscle strength and leg asymmetries in elite runners and cyclists: Original research article. International SportMed Journal, 15(3). hdl.handle.net/10520/EJC159075
ISSN: 2014-0983
Rebut: 16 d’octubre de 2025
Acceptat: 27 de febrer de 2026
Publicat: 1 de juliol de 2026
Editat per: © Generalitat de Catalunya Departament de la Presidència Institut Nacional d’Educació Física de Catalunya (INEFC)
© Copyright Generalitat de Catalunya (INEFC). Aquest article està disponible a la url https://www.revista-apunts.com/. Aquest treball està publicat sota una llicència Internacional de Creative Commons Reconeixement 4.0. Les imatges o qualsevol altre material de tercers d’aquest article estan incloses a la llicència Creative Commons de l’article, tret que s’indiqui el contrari a la línia de crèdit; si el material no s’inclou sota la llicència Creative Commons, els usuaris hauran d’obtenir el permís del titular de la llicència per reproduir el material. Per veure una còpia d’aquesta llicència, visiteu https://creativecommons.org/licenses/by/4.0/deed.ca


