Evaluación de las asimetrías de potencia, cinemática y oxigenación muscular entre piernas durante una carrera incremental de 400 m en triatletas
Jordi Montraveta
Ignacio Fernández-Jarillo
Xavier Iglesias

Diego Chaverri
*Correspondencia: Diego Chaverri dchaverri@gencat.cat
Citación
Montraveta, J., Fernández-Jarillo, I., Iglesias, X., & Chaverri, D. (2026). Assessing power, kinematic, and muscle oxygenation asymmetries between legs during incremental 400-m track running in triathletes. Apunts. Educación Física y Deportes, 165, 70-81. https://doi.org/10.5672/apunts.2014-0983.es.2026.165.07
Resumen
Correr sobre una pista de 400 m puede inducir asimetrías en la potencia desarrollada (PD), la cinemática o la saturación muscular de oxígeno (SmO₂). El objetivo principal de este estudio era determinar si correr sobre una pista de 400 m induce diferencias medibles en la potencia desarrollada, la cinemática y la saturación muscular de oxígeno entre la pierna interior y la exterior. El objetivo secundario era evaluar la magnitud de las asimetrías en la potencia desarrollada, la cinemática y la saturación muscular de oxígeno en triatletas durante la carrera en pista. Los datos cinemáticos y de PD se recogieron mediante sensores Stryd colocados en ambas zapatillas, mientras que los datos de SmO₂ se obtuvieron mediante dispositivos de espectroscopia de infrarrojos cercanos (NIRS, por sus siglas en inglés) colocados sobre el músculo vasto lateral (VL) de ambas piernas. Aunque la SmO₂, la PD y los parámetros cinemáticos mostraron cambios correspondientes al aumento de la velocidad durante el test de VAM-EVAL, un protocolo de carrera incremental en el que la velocidad aumenta a razón de 0.5 km·h-1 cada minuto hasta el agotamiento, el ANOVA de mediciones repetidas reveló efectos principales significativos de la pierna (interior frente a exterior) en la potencia desarrollada (PD), la longitud de zancada (LZ), el tiempo de contacto con el suelo (TCS) y la oscilación vertical (OV). Sin embargo, los análisis post hoc solo identificaron diferencias entre extremidades significativas en la PD al 80 % de la velocidad aeróbica máxima (VAM), y en la LZ al 70 % y al 80 % de la VAM. En cuanto a las asimetrías, el ANOVA de mediciones repetidas no mostró diferencias significativas entre intensidades, excepto en la SmO₂, con cambios significativos observados entre el 60 % y el 90 % y entre el 70 % y el 90 % de la VAM. Estas observaciones ponen de relieve el potencial de las asimetrías de SmO₂ medidas mediante NIRS para detectar de forma no invasiva diferencias en la utilización de oxígeno entre una pierna y la otra, especialmente a intensidades elevadas (90 % de la VAM). Esta tecnología podría ayudar a deportistas y entrenadores a identificar desequilibrios en el aporte y la utilización de oxígeno entre piernas, factor crítico para optimizar el rendimiento muscular.
Introducción
Correr sobre una pista de 400 m es fundamental para el entrenamiento y la evaluación del rendimiento, ya que proporciona un entorno controlado para valorar la resistencia y la eficiencia cinemática (Léger y Boucher, 1980). Estas pistas cuentan con dos rectas (de aproximadamente 84.39 m cada una) y dos curvas con un radio de alrededor de 36.5 m (World Athletics, 2019). Las curvas plantean desafíos biomecánicos específicos, ya que obligan a los corredores a gestionar giros continuos hacia la izquierda. Este movimiento asimétrico impone distintas demandas mecánicas y fisiológicas sobre la pierna interior y la exterior debido a las variaciones en la carga, con posibles consecuencias para el rendimiento y el riesgo de lesiones (Alt et al., 2015). En respaldo de esta perspectiva, Gilgen-Ammann et al. (2017) observaron que la asimetría de la marcha era significativamente mayor en corredores con antecedentes de lesión que en corredores sin lesiones, lo que sugiere que las asimetrías persistentes podrían reflejar vulnerabilidades subyacentes. Esto plantea la cuestión de si las asimetrías preexistentes podrían ser la causa de estas lesiones (Knapik et al., 1991; Vasquez-Bonilla et al., 2022, 2023).
Para comprender mejor el origen de estas asimetrías, es esencial analizar el modo en que la carrera en curva afecta a determinadas variables cinemáticas tales como la cadencia (CAD), la longitud de zancada ((LZ) y el tiempo de contacto con el suelo (TCS). En respuesta a las fuerzas centrífugas generadas durante el giro, los deportistas suelen presentar una LZ más corta y un TCS más largo en la pierna interior que en la exterior, especialmente a velocidades de carrera más altas. La pierna interior actúa como estabilizadora bajo mayores fuerzas compresivas, mientras que la exterior proporciona una mayor propulsión para dar respuesta a la demanda biomecánica de la curva (Chang y Kram, 2007). En el estudio de Chang y Kram (2007), se observaron asimetrías entre la pierna interior y la exterior durante esprints en una curva de 6 m de radio. La pierna exterior mostró una LZ significativamente mayor que la interior (1.70 ± 0.10 m frente a 1.53 ± 0.02 m; p < .05). Aunque también se observaron diferencias en la frecuencia de paso (FP) (3.88 ± 0.13 pasos s⁻¹ frente a 3.56 ± 0.12 pasos s⁻¹) y el TCS (0.203 ± 0.008 s frente a 0.190 ± 0.006 s), estas diferencias carecieron de significación estadística. Hamill et al. (1987) investigaron el modo en que los corredores ajustaban la función de las extremidades inferiores al recorrer las curvas de una pista de 400 m. Su estudio reveló asimetrías significativas entre las extremidades derecha e izquierda durante la carrera en curva, especialmente en la fase inicial del contacto del pie. En concreto, se observaron diferencias en los ángulos de toma de contacto, los ángulos máximos de pronación y las fuerzas de reacción del suelo mediolaterales. Estas observaciones sugieren que las transiciones entre las secciones curvas y rectas de la pista obligan a los corredores a adaptar dinámicamente sus patrones de carrera. Los giros continuos hacia la izquierda en las curvas imponen demandas mecánicas y fisiológicas distintas a cada pierna, lo que puede afectar a variables como la oscilación vertical (OV) y la CAD (Hamill et al., 1987).
Con el tiempo, estas cargas mecánicas asimétricas podrían contribuir al desarrollo de asimetrías entre extremidades, especialmente en deportistas que entrenan con frecuencia en pistas con curvas. Este concepto hace referencia a desequilibrios de rendimiento entre las extremidades opuestas del cuerpo (Bishop et al., 2017, 2022; Fox et al., 2023). Si bien las asimetrías entre extremidades se han estudiado tradicionalmente en deportes multidireccionales que implican acciones explosivas y cambios rápidos de dirección (Bishop et al., 2017; Loturco et al., 2019), algunos estudios recientes también han analizado estos desequilibrios en disciplinas lineales y de resistencia como la carrera y el triatlón (Helme et al., 2021; D’Hondt et al., 2024; Jacques et al., 2021). Estas observaciones sugieren que las asimetrías son relevantes no solo en los deportes de equipo, sino también en actividades caracterizadas por un movimiento continuo y repetitivo. Las asimetrías en la distribución de la masa magra, la fuerza muscular o la potencia desarrollada (PD) se han relacionado con problemas de rendimiento y con un mayor riesgo de lesiones (Bell et al., 2014; Knapik et al., 1991).
Las revisiones sistemáticas han subrayado la compleja relación entre la asimetría entre extremidades y el rendimiento deportivo. Por ejemplo, Bishop et al. (2017) concluyeron que las asimetrías entre extremidades observadas en un amplio abanico de tareas suelen tener un efecto negativo sobre el rendimiento físico. Sin embargo, sus conclusiones pusieron de relieve una variabilidad considerable, ya que algunos estudios notificaron una asociación mínima o nula entre la asimetría y los resultados de rendimiento. Del mismo modo, D’Hondt et al. (2024) señalaron una elevada heterogeneidad metodológica en los estudios sobre asimetría entre extremidades y carrera de resistencia, lo que dificulta extraer conclusiones definitivas. A pesar de que algunos indicadores de asimetría, como las asimetrías superiores al 10 % en el salto con contramovimiento unilateral (SCML) o las diferencias de fuerza entre extremidades, pueden afectar negativamente al rendimiento en carrera (Bishop et al., 2017), los datos científicos no siempre son coherentes, lo que subraya la necesidad de estudios de calidad para determinar umbrales y consecuencias prácticas para el entrenamiento.
En el triatlón, las asimetrías cinemáticas podrían verse más influidas por las demandas mecánicas prolongadas y repetitivas del tramo en bicicleta que por la propia transición. La fatiga acumulada durante el tramo en bicicleta puede alterar el control neuromuscular durante la carrera posterior, lo cual podría aumentar las asimetrías entre extremidades (Millet y Vleck, 2000). Heiden y Burnett (2003) demostraron que los patrones de activación muscular en las piernas durante la carrera se alteran significativamente después del tramo en bicicleta, con cambios notables en músculos importantes de las extremidades inferiores como el bíceps femoral y el vasto lateral. Del mismo modo, Connick y Li (2015) notificaron un aumento de la variabilidad del tiempo de zancada y una reducción de la longitud de zancada durante la carrera posterior al tramo en bicicleta, lo que indica una alteración del control locomotor probablemente vinculada a la fatiga previa. Olcina et al. (2019) observaron una disminución de la longitud de zancada después del tramo en bicicleta, lo que apunta a una alteración de la activación neuromuscular como factor contribuyente.
Además de las variables cinemáticas y de activación muscular, los parámetros de fisiología muscular también son relevantes para comprender mejor las asimetrías entre extremidades. La saturación muscular de oxígeno (SmO₂) proporciona una información valiosa para evaluar posibles diferencias entre extremidades. Una disminución de la SmO₂ refleja un mayor desequilibrio entre el aporte y la utilización de oxígeno, lo que indica una mayor extracción muscular de oxígeno durante el ejercicio (van der Zwaard et al., 2016). Mediante espectroscopia de infrarrojos cercanos (NIRS), los investigadores han podido evaluar si existe variabilidad o asimetrías en la utilización de oxígeno entre una pierna y la otra durante actividades como el ciclismo (Sendra-Pérez et al., 2025; Skotzke et al., 2024). Olcina et al. (2019) analizaron el efecto del tramo en bicicleta previo sobre el rendimiento en carrera, la longitud de zancada y la saturación muscular de oxígeno (SmO₂) en triatletas, y sus resultados indicaron que el tramo en bicicleta de alta intensidad antes de la carrera puede perjudicar el rendimiento al reducir la longitud de zancada y limitar la utilización periférica de oxígeno en músculos que presentan una concentración elevada de SmO₂. Estas observaciones sugieren que la fatiga previa inducida por el esfuerzo en bicicleta no solo afecta a la eficiencia biomecánica, sino que también genera desequilibrios localizados de oxigenación, lo que podría agravar las asimetrías musculares durante el tramo de carrera posterior. Se han notificado observaciones similares en otros deportes como el fútbol y el rugby, en los que las asimetrías de SmO₂ se correlacionan con una menor eficiencia en el rendimiento y un mayor riesgo de lesiones (Vasquez-Bonilla et al., 2022, 2023).
Actualmente, la tecnología portátil permite hacer un seguimiento en tiempo real de las variables cinemáticas, la PD y la SmO₂ para evaluar las asimetrías. Dispositivos como el sensor Stryd proporcionan datos detallados sobre la PD, la LZ el TCS y la OV, que facilitan el análisis del rendimiento (van Rassel et al., 2023). Estas herramientas ayudan a deportistas y entrenadores a identificar diferencias entre extremidades en la producción mecánica y la SmO₂, lo que permite orientar el entrenamiento para optimizar el rendimiento y reducir el riesgo de lesiones. En estudios previos, se ha demostrado su utilidad en este contexto. Por ejemplo, Bini y Hume (2015) hallaron asimetrías notables en la fuerza aplicada al pedal durante pruebas contrarreloj de ciclismo, mientras que Yanci (2014) notificó asimetrías de PD entre la pierna dominante y la no dominante en deportistas de resistencia. Ambos estudios sugieren que abordar estos desequilibrios podría mejorar el rendimiento y reducir el riesgo de lesiones.
Sobre esta base, la asimetría entre extremidades se define como el desequilibrio o diferencia entre las extremidades inferiores que puede manifestarse en dimensiones funcionales, cinemáticas o cinéticas. En corredores de resistencia, la investigación ha notificado asimetrías funcionales de aproximadamente el 16-17 % y diferencias biomecánicas que oscilan entre el 3 % y el 54 %, según la tarea y el indicador que se emplee (D’Hondt et al., 2024). Si bien los datos que cuantifican específicamente su prevalencia general en triatletas siguen siendo escasos debido a la falta de estudios previos en esta población, se han documentado diferencias laterales significativas en características neuromusculares. En concreto, se ha demostrado que correr después de pedalear altera los patrones de activación muscular (por ejemplo, reduce la activación del sóleo en una extremidad), pese a no observarse diferencias en la cinética o la cinemática externas (Jacques et al., 2021). Dadas las demandas biomecánicas de la carrera en pista y la presencia documentada de asimetrías entre extremidades en deportistas de resistencia y triatletas, el presente estudio se planteó los siguientes objetivos:
Analizar si correr sobre una pista de 400 m induce diferencias medibles en la potencia, la cinemática y la saturación muscular de oxígeno entre la pierna interior y la exterior.
Evaluar la magnitud de las asimetrías de potencia, cinemática y saturación muscular de oxígeno en triatletas durante la carrera en pista.
Materiales y métodos
Participantes
Catorce triatletas varones de nivel nacional participaron voluntariamente en este estudio (Tabla 1). Los criterios de inclusión fueron estar federado y competir activamente en pruebas nacionales de triatlón. Los criterios de exclusión fueron padecer algún trastorno cardíaco, presentar una lesión en ese momento o haberla sufrido en los dos meses anteriores. Además, se excluyó a las personas con un grosor del tejido adiposo (GTA) superior a 7 mm para minimizar la interferencia con la calidad de la señal NIRS, ya que un GTA mayor puede atenuar la luz infrarroja cercana y afectar a la precisión de las lecturas de SmO₂ (McManus et al., 2018). El GTA se calculó como 0.5 × el grosor medio del pliegue cutáneo. Todos los participantes dieron su consentimiento informado por escrito, y el protocolo del estudio fue aprobado por el Comité de Ética de Investigación Clínica de la Administración Deportiva Catalana (026/CEICGC/2023).

Materiales
Los parámetros cinemáticos y de PD se midieron mediante el sensor Stryd (Stryd Inc., Boulder, CO, EE. UU.), que se ha validado previamente para la estimación de la potencia en carrera en condiciones controladas (Cerezuela-Espejo et al., 2021). Los sensores Stryd se fijaron a los cordones de cada zapatilla, colocados en posición central sobre el dorso del pie, de acuerdo con las instrucciones del fabricante. El dispositivo obtuvo muestras y exportó los datos procesados a 1 Hz, mientras que los datos inerciales internos se registraron a frecuencias más altas (hasta ~100 Hz). La SmO₂ se midió con un dispositivo NIRS de onda continua (MOXY, Hutchinson, Minnesota, Estados Unidos) utilizando cuatro longitudes de onda (680, 720, 760 y 800 nm). La SmO2 se midió en forma de porcentaje aplicando la siguiente ecuación: SmO₂ = [oxihemoglobina (O2Hb) / (oxihemoglobina (O2Hb) + desoxihemoglobina (HHb))] * 100, donde O₂Hb representa la oxihemoglobina y HHb representa la desoxihemoglobina. El dispositivo MOXY es un sistema NIRS portátil y no invasivo diseñado para evaluaciones de campo de la oxigenación muscular. Su validez y fiabilidad para medir la SmO₂ durante el ejercicio se han demostrado previamente (Feldmann et al., 2019). El dispositivo NIRS se colocó sobre la zona central del músculo vasto lateral (VL) en ambas piernas, a medio camino entre el trocánter mayor y el epicóndilo femoral lateral (McManus et al., 2018). Los sensores se fijaron con cinta adhesiva (Hypafix; BSN Medical, Hamburgo, Alemania). La frecuencia de muestreo se dejó en el modo predeterminado del dispositivo: los datos se actualizaban cada dos segundos y se suavizaban tomando como referencia los 10 segundos anteriores.
Procedimientos
La colocación de los sensores Moxy se determinó con una cinta métrica y un rotulador. Además, el grosor del pliegue cutáneo se midió en el lugar donde se colocó el dispositivo NIRS mediante un plicómetro Harpenden (precisión: 0.20 mm). A continuación, se fijaron los sensores Stryd a ambas zapatillas.
Todos los participantes realizaron un test de VAM-EVAL (García y Secchi, 2013) sobre una pista de 400 m.Según el protocolo original, no se hizo un calentamiento estandarizado antes de la prueba, ya que la baja velocidad inicial de carrera proporcionaba un calentamiento progresivo.La velocidad de carrera se controló mediante una señal acústica. La velocidad inicial se estableció en 8.5 km·h⁻¹ y aumentó a razón de 0.5 km·h⁻¹ por minuto. Se colocaron conos cada 20 m para ayudar a los participantes a ajustar su ritmo a las señales acústicas. La prueba finalizó cuando los participantes se detuvieron por agotamiento o no alcanzaron el cono marcado en dos ocasiones consecutivas. La velocidad aeróbica máxima (VAM) se definió como la velocidad alcanzada durante la última etapa completada; no se tuvieron en cuenta las etapas incompletas (García y Secchi, 2013).
Las sesiones de prueba se realizaron en una pista de atletismo al aire libre en Barcelona entre el 27 de febrero y el 23 de abril de 2024, en condiciones ambientales medias de 21.1 ± 5.7 °C y 74.6 ± 16.6 % de humedad relativa. Se indicó a los participantes que se abstuvieran de realizar entrenamientos de alto volumen o alta intensidad durante las 24 horas previas a la prueba
Análisis estadístico
Los datos cinemáticos y de SmO₂ se filtraron cada 2 segundos antes del análisis y se alinearon temporalmente con el tiempo cero (t = 0) para su sincronización. El tiempo cero se definió como el inicio del test de VAM-EVAL, identificado por el primer aumento de la velocidad de carrera indicado por la señal acústica, y se sincronizó entre dispositivos utilizando las marcas temporales internas de los sistemas de registro. Se calcularon datos estadísticos descriptivos (media ± desviación típica [DT]) para todas las variables.
Los niveles de intensidad (60 %, 70 %, 80 % y 90 % de la velocidad aeróbica máxima [VAM]) se determinaron a partir de los resultados del test de VAM-EVAL. Los valores medios de ambas piernas se obtuvieron a partir de los 20 segundos centrales de cada intervalo de un minuto en cada nivel de intensidad.
La normalidad se evaluó mediante la prueba de Shapiro-Wilk. Con la excepción de algunos índices de asimetría PD ASIM, CAD ASIM, OV ASIM, TCS ASIM, y LZ ASIM), todas las variables cumplieron el supuesto de normalidad. En consecuencia, se realizó un análisis de la varianza (ANOVA) de mediciones repetidas de dos factores para comparar las diferencias entre piernas (pierna interior frente a pierna exterior) y niveles de intensidad (60 %, 70 %, 80 % y 90 % de la VAM) en la PD, los parámetros cinemáticos (CAD, OV, TCS y LZ) y la SmO₂.
Se realizó un ANOVA unifactorial independiente para examinar las diferencias en los índices de asimetría de la PD, los parámetros cinemáticos (CAD, OV, TCS y LZ) y la SmO₂entre niveles de intensidad. La asimetría (%) de cada variable se calculó de acuerdo con la siguiente ecuación:
donde Xr representa la SmO2 registrada en la pierna derecha y Xl representa la SmO2 correspondiente registrada en la pierna izquierda. Este enfoque sigue protocolos establecidos (Karamanidis et al., 2003; Knapik et al., 1991).
Tanto en el ANOVA de mediciones repetidas de dos factores como en el ANOVA unifactorial de los índices de asimetría, se realizaron comparaciones por pares post hoc utilizando el método de corrección de Holm para controlar la tasa de error familiar debida a las comparaciones múltiples, y se notifican los valores de p ajustados. Para las comparaciones por pares posteriores al ANOVA de dos factores, los tamaños del efecto se calcularon mediante la d de Cohen, con valores de 0.2, 0.5 y 0.8 interpretados como efectos pequeños, medios y grandes, respectivamente (Cohen, 1988). Para el ANOVA unifactorial aplicado a los índices de asimetría con distribución normal (p. ej., SmO₂ ASIM), los tamaños del efecto se estimaron mediante eta al cuadrado parcial (η²p), con umbrales de .01, .06 y .14 considerados efectos pequeños, medios y grandes, respectivamente (Richardson, 2011). En cambio, para los índices de asimetría que no cumplieron el supuesto de normalidad (p. ej., PD ASIM, CAD ASIM, OV ASIM, TCS ASIM y LZ ASIM), se aplicó la prueba de Friedman, seguida de pruebas de rangos con signo de Wilcoxon y corrección de Holm para las comparaciones por pares; en estos casos, los tamaños del efecto se calcularon mediante la W de Kendall, interpretada como niveles de concordancia pequeños (.1), moderados (.3) y grandes (.5) (Tomczak y Tomczak, 2014).
Todos los análisis estadísticos se realizaron con Microsoft Excel (versión 16.81, 24011420) y JASP (versión 0.18.3). La significación estadística se fijó en p < .05 para los efectos globales.
Resultados
Los participantes (n = 14) alcanzaron una frecuencia cardíaca máxima (FCmáx) de 183.5 ± 15.6 lpm y una VAM de 18.53 ± 1.02 km·h-1 durante el test de VAM-EVAL. La SmO2, la PD y los parámetros cinemáticos mostraron cambios progresivos en respuesta al aumento de la velocidad de carrera a lo largo de la prueba.
Durante las etapas incrementales del test de VAM-EVAL (60 %, 70 %, 80 % y 90 % de la VAM), la PD, la CAD y la LZ aumentaron significativamente en ambas piernas en todas las intensidades, mientras que el TCS disminuyó significativamente en todas las intensidades (todas las p inferiores a .05) (Tabla 2). Se observó un efecto principal significativo de la variable Pierna en la PD (F(1,13) = 13.62, p = .003, η²p = .512), la OV (F(1,13) = 7.97, p = .014, η²p = .38), el TCS (F(1,13) = 6.51, p = .024, η²p = .334) y la LZ (F(1,13) = 21.26, p < .001, η²p = .621). Sin embargo, las comparaciones post hoc entre la pierna interior y la exterior en cada nivel de intensidad solo alcanzaron significación estadística tras la corrección de Holm en la PD al 80 % de la VAM (t(13) = −4.455, p = .003, d = −0.29), la LZ al 70 % de la VAM (t(13) = −6.298, p < .001, d = −0.37)) y la LZ al 80 % de la VAM (t(13) = −4.717, p < .001, d = −0.47) ). Esto sugiere un efecto principal constante de la variable Pierna en la OV y el TCS, con diferencias sistemáticas de una pierna respecto de la otra en todas las intensidades. Sin embargo, la ausencia de una interacción significativa entre Pierna e Intensidad indica que la magnitud de esta diferencia entre extremidades se mantuvo estable, independientemente del nivel de intensidad. No se hallaron diferencias significativas entre piernas en la CAD (p = .313, η²p = .08) ni en la SmO₂ (p = .976, η²p = .00).
Tabla 2
Datos descriptivos de ambas piernas en PD, CAD, OV, TCS, LZ y SmO₂ durante el test de VAM-EVAL (n = 14; se indican las diferencias significativas (p < .05) entre niveles de intensidad)
El análisis del tamaño del efecto reveló diferencias entre extremidades entre moderadas y grandes en la PD, la OV, el TCS y la LZ (η²p entre .33 y .62). En cambio, el TCS, la LZ y la PD mostraron adaptaciones intramiembro marcadas con el aumento de la intensidad, tal y como reflejan los significativos efectos de intensidad y los valores de eta al cuadrado parcial superiores a .95 (Tabla 3). Tanto la pierna interior como la exterior mostraron tamaños del efecto intramiembro muy grandes en respuesta al aumento de la intensidad, especialmente en el TCS, la LZ y la PD. Los tamaños del efecto oscilaron entre 1.07 y 3.26 en la PD, entre 1.75 y 6.97 en el TCS, y entre 1.38 y 4.76 en la LZ La CAD y la SmO₂ también mostraron cambios intramiembro, aunque de menor magnitud. Los tamaños del efecto oscilaron entre 0.52 y 2.16 en la CAD y entre 0.08 y 1.15 en la SmO₂, lo que sugiere ajustes intramiembro entre moderados y grandes, especialmente en los niveles de intensidad más altos. En cambio, la OV solo mostró tamaños del efecto pequeños e inconsistentes (d = 0.08 a 0.39), lo que indica una respuesta limitada al aumento de la intensidad.
Tabla 3
Resultados del ANOVA de mediciones repetidas de dos factores para PD, CAD, OV, TCS, LZ y SmO₂ durante el test de VAM-EVAL (n = 14)
En cuanto a los valores de asimetría entre extremidades, no se observaron diferencias significativas en la PD, la CAD, la LZ, la OV ni el TCS entre los distintos niveles de intensidad (Tabla 4). Sin embargo, la asimetría en la SmO₂ mostró diferencias significativas entre el 60 % y el 90 % y entre el 70 % y el 90 % de la VAM (p < .05).
Tabla 4
Datos descriptivos de los índices de asimetría de PD, CAD, OV, TCS, LZ y SmO₂ entre niveles de intensidad (60 %, 70 %, 80 % y 90 % de la VAM; se indican las diferencias significativas entre niveles de intensidad [p < .05])
Tabla 5
Resultados de los análisis estadísticos de los índices de asimetría entre extremidades entre niveles de intensidad durante el test de VAM-EVAL
El análisis del tamaño del efecto reveló efectos entre pequeños y grandes en la asimetría de la SmO₂ entre intensidades, que oscilaron entre pequeños (60-70 % de la VAM: d = 0.081; 70-80 % de la VAM: d = 0.266), moderados (60-80 % de la VAM: d = 0.347; 80-90 % de la VAM: d = 0.574) y grandes (60-90 % de la VAM: d = 0.921; 70-90 % de la VAM: d = 0.840) (Tabla 4). Estos resultados sugieren que la asimetría de SmO₂ aumenta progresivamente a medida que se incrementa la intensidad de carrera.
La Figura 1 ilustra los patrones de SmO₂ de ambas piernas en dos participantes representativos. El panel izquierdo muestra un participante con asimetrías marcadas, mientras que el derecho muestra uno sin asimetría observable. En el participante asimétrico, aunque los valores absolutos de SmO₂ difieren considerablemente entre piernas, los perfiles temporales de saturación siguen siendo notablemente similares entre intensidades.
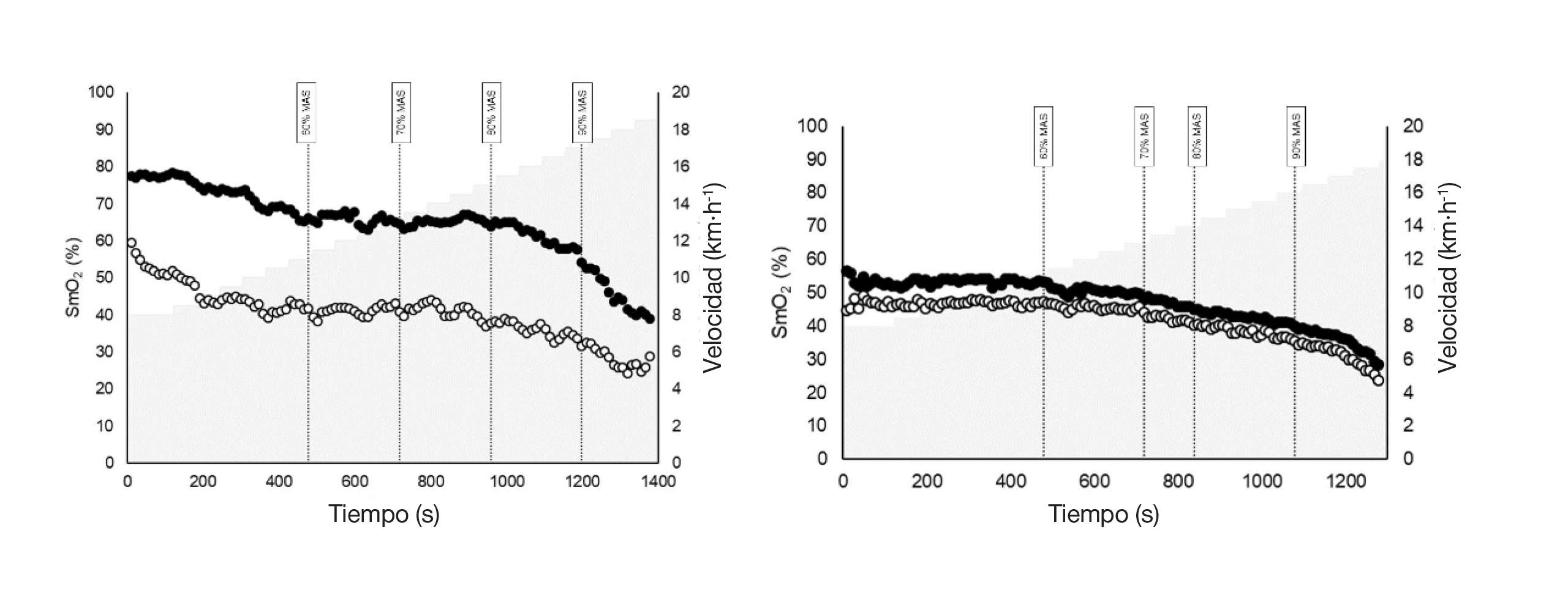
Nota. Los valores de SmO₂ se expresan en forma de porcentaje (%), y la velocidad de carrera se expresa en kilómetros por hora (km·h-1). La línea negra representa la pierna exterior, mientras que la línea de color claro representa la pierna interior. VAM = velocidad aeróbica máxima.
Discusión
El presente estudio es uno de los primeros en analizar las asimetrías de potencia, cinemática y oxigenación muscular entre la pierna interior y la exterior durante una carrera incremental en pista de 400 m en un entorno realista al aire libre. La mayoría de los estudios previos de este ámbito se han realizado en condiciones controladas de laboratorio en interiores, que tal vez no capten plenamente la complejidad biomecánica y fisiológica de la carrera en condiciones reales.
El primer objetivo era determinar si correr sobre una pista de 400 m induce diferencias medibles en la PD, las variables cinemáticas (CAD, OV, TCS, LZ) y la SmO₂ entre la pierna interior y la exterior. Las principales observaciones revelaron efectos principales significativos de la variable Pierna en la PD, la LZ, la OV y el TCS, lo que indica diferencias entre extremidades sistemáticas en todos los niveles de intensidad. Sin embargo, los análisis post hoc solamente identificaron diferencias con significación estadística en determinadas intensidades (PD al 80 % de la VAM, y LZ al 70 % y al 80 % de la VAM), mientras que no se observaron diferencias entre extremidades significativas en la CAD ni en la SmO₂. Además, la ausencia de interacciones significativas entre Pierna e Intensidad sugiere que estas asimetrías se mantuvieron estables a lo largo del protocolo incremental, en lugar de aumentar con la intensidad del ejercicio.
El segundo objetivo fue evaluar la magnitud y la progresión de las asimetrías entre extremidades a medida que aumentaba la intensidad de carrera. Los principales hallazgos mostraron que los índices de asimetría de la PD, la CAD, la OV, el TCS y la LZ no cambiaron significativamente entre los niveles de intensidad, lo que sugiere una simetría mecánica estable a lo largo de la prueba. En cambio, la asimetría de SmO₂ aumentó progresivamente con la intensidad, con tamaños del efecto grandes observados al 90 % de la VAM. Esta divergencia fisiológica sugiere un desequilibrio creciente en la oxigenación muscular local entre extremidades bajo un mayor estrés metabólico; esto posiciona la SmO₂ como un marcador sensible y potencialmente precoz de asimetría dependiente de la carga durante la carrera a alta velocidad.
Estas observaciones coinciden parcialmente con estudios previos que notificaron alteraciones cinemáticas durante la carrera en curva; sin embargo, a la hora de comparar los resultados deben tenerse en cuenta algunas diferencias metodológicas importantes. Por ejemplo, los estudios de Chang y Kram (2007) y Hamill et al. (1987) demostraron que la carrera en curva afecta a variables cinemáticas relevantes como la CAD, la LZ, la FP, la OV y el TCS. Es importante reconocer las considerables diferencias entre sus condiciones experimentales y las del presente estudio. En concreto, Chang y Kram (2007) investigaron la carrera en curvas con un radio mucho menor (6 m) que el de una pista de atletismo estándar (36.5 m), y sus participantes corrieron a velocidades máximas de esprint, a diferencia de las intensidades submáximas utilizadas en nuestro protocolo (hasta el 100 % de la VAM; 5.15 ± 0.28 m·s-1). Del mismo modo, Hamill et al. (1987) examinaron la carrera a una velocidad considerablemente superior (6.31 ± 5 m·s-1) al ritmo mantenido por los triatletas en nuestro estudio. Estas diferencias metodológicas produjeron fuerzas centrífugas mucho mayores, lo que amplificó las disparidades cinemáticas que constataron entre la pierna interior y la exterior.
También observamos que, a medida que aumentaba la intensidad de carrera, se producían incrementos significativos en la PD, la CAD y la LZ en ambas piernas en todos los niveles de intensidad. En concreto, la PD en la PI aumentó un 13.6 %, un 13.2 % y un 10.4 % entre el 60 % y el 70 %, entre el 70 % y el 80 %, y entre el 80 % y el 90 % de la VAM, respectivamente. De forma similar, la PD en la PE aumentó un 13.9 %, un 11.2 % y un 11.9 % en los mismos intervalos. En cuanto a la CAD, ambas piernas mostraron incrementos moderados pero constantes con la intensidad. La CAD en la PI aumentó un 2.1 %, un 2.2 % y un 3.4 %, mientras que la PE mostró incrementos ligeramente menores pero progresivos del 1.7 %, el 2.4 % y el 3.6 % en los intervalos correspondientes. La LZ también aumentó con la intensidad, aunque la tasa de incremento disminuyó al acercarse al 90 % de la VAM. En la PI, la LZ aumentó un 13.7 %, un 11.5 % y un 8.0 %, mientras que en la PE se observaron incrementos del 13.2 %, el 11.1 % y el 9.3 %. Por el contrario, el TCS disminuyó de forma constante en ambas piernas a medida que aumentó la intensidad. En la PI, el TCS se redujo un 9.8 %, un 8.6 % y un 6.8 % entre el 60 % y el 70 %, entre el 70 % y el 80 %, y entre el 80 % y el 90 % de la VAM, respectivamente. En la PE, las reducciones fueron del 9.8 %, el 8.9 % y el 7.6 % en los mismos intervalos de intensidad. Estas observaciones coinciden con las de Patoz et al. (2023), especialmente en cuanto a la coherencia de las respuestas biomecánicas a medida que aumentan las intensidades de carrera. Al igual que en sus observaciones al 90 %-120 % de la velocidad aeróbica pico, nuestros datos no revelaron interacciones significativas entre Pierna e Intensidad (p > .05), lo que indica un patrón de adaptación uniforme en ambas extremidades. Además, ambos estudios notificaron una cadencia (frecuencia de zancada) y una oscilación vertical estables pese al aumento de la fatiga, lo que sugiere que los corredores priorizan el mantenimiento del ritmo de movimiento y el control del centro de masas. El efecto significativo de la intensidad observado en nuestros datos de tiempo de contacto con el suelo también respalda su interpretación de que los corredores adoptan estrategias espaciotemporales, como la modulación del tiempo de contacto, para optimizar la distribución de la carga durante esfuerzos de alta intensidad. Si bien ambos estudios se centraron en las respuestas biomecánicas a intensidades submáximas y cercanas a la máxima, nuestro protocolo fue ligeramente menos exigente. En nuestro estudio, los deportistas realizaron una prueba incremental continua sin periodos de descanso, mientras que Patoz et al. (2023) llevaron a cabo cuatro ensayos exhaustivos independientes a porcentajes fijos de la velocidad pico de cada participante, efectuados en orden aleatorizado.
Además de los ajustes mecánicos observados en los parámetros espaciotemporales, la respuesta fisiológica muscular también mostró tendencias relevantes a medida que aumentó la intensidad. En concreto, la saturación muscular de oxígeno (SmO₂) disminuyó progresivamente tanto en la pierna interior como en la exterior a lo largo del protocolo incremental. Esta respuesta encaja con estudios previos sobre oxigenación muscular durante ejercicio graduado, que notificaron una disminución continua de la SmO₂ debido al aumento de la demanda de extracción de oxígeno (Grassi et al., 1999). Las presentes observaciones confirman este patrón, especialmente a intensidades más altas, como el 90 % de la VAM.
No se observaron diferencias significativas entre la pierna interior y la exterior (p = .976), ni hubo una interacción significativa entre Pierna e Intensidad (p = .068). Sin embargo, se observó un fuerte efecto principal de la intensidad (p < .001, η²p = .697), lo que indica que la SmO₂ respondió de forma marcada a los aumentos de la velocidad de carrera. Estos resultados sugieren que los cambios observados en la SmO₂ reflejan una adaptación fisiológica global al aumento de la demanda metabólica, sin una evidencia clara de desequilibrios funcionales entre la pierna interior y la exterior en cuanto a la oxigenación muscular.
El objetivo secundario de este estudio era evaluar la magnitud de las asimetrías de la PD, las variables cinemáticas (CAD, OV, TCS y LZ) y la SmO2 en triatletas de nivel nacional durante un test de VAM-EVAL. Las principales observaciones no mostraron diferencias significativas entre intensidades, excepto en la SmO2, que presentó cambios significativos entre el 60 % y el 90 % y entre el 70 % y el 90 % de la VAM. Estos resultados sugieren que la asimetría de SmO₂ es proporcional a la intensidad del ejercicio, especialmente al 90 % de la VAM, lo cual indica que, a medida que aumenta la intensidad de carrera, las diferencias en la utilización de oxígeno entre una pierna y la otra se hacen más evidentes en los triatletas.
Para contextualizar mejor estas conclusiones, es esencial considerar los mecanismos fisiológicos subyacentes que podrían explicar las asimetrías observadas. La presencia de una asimetría significativa de SmO₂, pese a una producción mecánica simétrica en parámetros como la potencia y la longitud de zancada, apunta al papel de factores musculares intrínsecos. En particular, las variaciones locales en la capilarización y las adaptaciones vasculares podrían contribuir a estas disparidades. La capilarización es un factor determinante del aporte y la difusión de oxígeno; concretamente, parámetros como la densidad capilar y el número de contactos capilares por área de fibra (CC/AF) se asocian con una extracción de O₂ más eficiente; esta relación es especialmente sólida entre los jóvenes (Chilibeck et al., 1997). Asimismo, las adaptaciones vasculares provocadas por el entrenamiento de resistencia suelen ser localizadas, con una mayor respuesta microvascular principalmente en los tejidos directamente implicados en el esfuerzo (Soares et al., 2018).
Los procesos fisiológicos internos pueden seguir siendo asimétricos debido a diferencias en la perfusión muscular local o el metabolismo oxidativo. En este contexto, determinados mecanismos reflejos del sistema nervioso autónomo, como el reflejo presor del ejercicio, pueden aumentar el riego sanguíneo de los músculos que realizan un mayor esfuerzo, lo que podría reducir la disponibilidad de oxígeno en otros músculos menos implicados en la tarea o con una función auxiliar. Estos mecanismos forman parte de la respuesta autónoma general al aumento de la demanda metabólica y participan en la modulación de las respuestas vasculares específicas de cada extremidad durante el ejercicio intenso (Orcioli-Silva et al., 2024).
Además, estrategias neuromusculares como la coactivación muscular contribuyen a estas asimetrías internas. A velocidades de carrera elevadas, la coactivación global aumenta y se produce antes y durante periodos más breves para incrementar la rigidez de toda la extremidad y la estabilización articular (Fiori et al., 2024). Cabe destacar que la relación entre excitación muscular (RMS) y SmO₂ no es uniforme en todos los músculos. Mientras que los músculos generadores de potencia, como el vasto lateral, muestran un patrón inverso claro (la RMS aumenta mientras la SmO₂ disminuye) debido a una mayor demanda metabólica, algunos estabilizadores, como el gastrocnemio medial, mantienen un nivel estable de SmO₂ y de RMS durante todo el ejercicio (Sendra-Pérez et al., 2025). Esto refleja sus funciones diferenciadas, ya que los músculos estabilizadores participan más en la regulación de la carga que en la generación de fuerza. Estas observaciones ponen de relieve la compleja interacción entre mecanismos neuromusculares y vasculares que modulan la dinámica de la SmO₂. Dado que el presente estudio se centró exclusivamente en el vasto lateral, el análisis de otros músculos con funciones diversas permitiría conocer de forma más completa estas asimetrías fisiológicas.
A pesar de que la mayoría de los estudios anteriores ha evaluado la simetría de SmO₂ mediante métodos estadísticos de concordancia y diferencias entre extremidades absolutas, nuestro análisis empleó un índice de asimetría normalizado (ASI%) para expresar la disparidad relativa entre piernas. En condiciones controladas de laboratorio, Sendra-Pérez et al. (2025) definieron la simetría a partir de diferencias absolutas de SmO₂ de entre el 10 % y el 20 %, mientras que Skotzke et al. (2024) utilizaron el análisis de Bland-Altman para establecer límites de concordancia de ± 20 %, y concluyeron que estas diferencias entre extremidades son frecuentes y no están vinculadas a la dominancia de la pierna. En cambio, nuestros valores de ASI% durante la carrera al aire libre oscilaron entre el 24.4 % y el 53.5 %, por encima tanto de los umbrales de laboratorio como del cambio mínimo detectable (CMD) del 18 % notificado por Skotzke et al. (2024) para la SmO₂ en el ciclismo.
Sin embargo, es importante señalar que el ASI% se ve influido por los valores absolutos bajos. Dado que los niveles de SmO₂ descendieron de forma marcada a intensidades elevadas (p. ej., ~25 % al 90 % de la VAM), incluso diferencias absolutas moderadas pueden generar valores de ASI% desproporcionadamente altos. Para validar nuestras observaciones, realizamos un análisis de Bland-Altman en todas las intensidades. Los resultados revelaron un sesgo sistemático mínimo al 60 %, 70 % y 80 % de la VAM (−2.6 % ± 24.0 %, +0.2 % ± 24.3 % y +0.3 % ± 28.3 %, respectivamente), con límites de concordancia relativamente coherentes. Al 90 % de la VAM, a pesar de que los valores de SmO₂ fueron más bajos, el sesgo siguió siendo pequeño (+1.7 %) y los límites de concordancia (± 25.68 %) no se ampliaron de forma significativa en comparación con las etapas anteriores. En comparación con Skotzke et al. (2024), que notificaron límites de concordancia de ± 20 % durante el esfuerzo estable en bicicleta, nuestros resultados muestran niveles de asimetría fisiológica ligeramente más amplios, pero todavía comparables en condiciones de carrera sobre el terreno más exigentes. Estos resultados sugieren que, si bien las asimetrías fisiológicas de SmO₂, tal como las capta el ASI%, sí se hacen más evidentes con el aumento de la intensidad, la magnitud de la asimetría relativa debe interpretarse con cautela, especialmente en condiciones de baja saturación de oxígeno.
Según la reciente revisión sistemática de D’Hondt et al. (2024), la relación entre la asimetría entre extremidades y el rendimiento en carrera de resistencia es compleja. Sus observaciones indican que algunos indicadores de asimetría funcionales, morfológicos, cinemáticos y cinéticos, aunque no todos, son inversamente proporcionales al rendimiento en carrera o no guardan con él una relación significativa. En el presente estudio, pese a mostrar asimetrías significativas de SmO₂, especialmente a intensidades elevadas, los triatletas alcanzaron un nivel de rendimiento de 3 según la clasificación propuesta por De Pauw et al. (2013). Para analizar las asimetrías en mayor profundidad, habría sido beneficioso realizar una evaluación preliminar de la composición corporal mediante absorciometría de rayos X de doble energía (DXA) para evaluar la composición corporal total e identificar posibles diferencias de masa muscular entre piernas. Branski et al. (2010) demostraron que la asimetría de masa magra influye en la asimetría de fuerza y potencia durante tareas de salto en deportistas universitarios. Asimismo, la realización de pruebas específicas de fuerza unilateral para corredores podría haber proporcionado información sobre posibles desequilibrios de fuerza entre extremidades. Este planteamiento habría permitido una interpretación más completa de las diferencias observadas en la saturación muscular de oxígeno.
Limitaciones y futuros estudios
Este estudio presenta algunas limitaciones que deben tenerse en cuenta. En primer lugar, el tamaño de la muestra fue relativamente reducido e incluyó únicamente triatletas varones de nivel nacional, lo cual podría limitar la generalización de las observaciones a otras poblaciones, tales como corredoras profesionales o personas que practican este deporte de manera informal. En segundo lugar, la SmO2 se midió exclusivamente en el vasto lateral. La inclusión de otros músculos implicados en la carrera podría haber permitido caracterizar de una forma más completa las asimetrías fisiológicas entre extremidades. En tercer lugar, no se incluyeron medidas directas de fuerza muscular unilateral, función neuromuscular o asimetría de la composición corporal, lo que podría haber contribuido a una interpretación más profunda de las asimetrías de SmO₂ observadas. Por último, aunque el hecho de realizar la prueba al aire libre acerca la evaluación a las condiciones reales de carrera, también podría haber introducido más variabilidad que un entorno de laboratorio.
Los futuros estudios deberían seguir haciendo hincapié en entornos realistas para reflejar con mayor precisión las demandas de la carrera al aire libre. Asimismo, la integración de análisis de composición corporal y evaluaciones específicas de fuerza de ambas piernas podría ayudar a identificar los orígenes fisiológicos de las asimetrías de SmO₂. La ampliación de estos estudios contribuiría a conocer mejor los mecanismos fisiológicos y biomecánicos subyacentes a los desequilibrios entre extremidades y sus posibles consecuencias para el rendimiento deportivo. Por último, los futuros estudios deberían incorporar herramientas de observación sistemática y análisis con vídeo (Soto et al., 2019) para aumentar la fiabilidad del análisis del movimiento en entornos deportivos aplicados.
Conclusiones
Las conclusiones de este estudio en triatletas varones de nivel nacional que realizaron una prueba incremental de carrera sobre pista de 400 m indican que, aunque no se observaron asimetrías en la potencia desarrollada ni en los parámetros cinemáticos, las asimetrías de saturación muscular de oxígeno se hicieron evidentes a medida que aumentó la intensidad de carrera. Estas asimetrías no fueron atribuibles a la trayectoria de carrera en curva, ya que no se hallaron diferencias significativas entre la pierna interior y la exterior, excepto en la potencia de salida al 80 % de la velocidad aeróbica máxima (VAM) y la longitud de zancada al 70 % y al 80 % de la VAM. Si bien los parámetros mecánicos se mantuvieron en gran medida simétricos, la divergencia progresiva en los valores de SmO₂ apunta a un desequilibrio fisiológico que se acentúa a intensidades más altas, lo cual destaca la SmO₂ como marcador sensible de asimetrías internas dependientes de la carga, incluso en ausencia de discrepancias mecánicas.
Agradecimientos
Los autores desean agradecer al Instituto Nacional de Educación Física de Cataluña y al Grupo de Investigación en Ciencias del Deporte INEFC Barcelona (GRCEIB, https://inefc-grceib.cat) su apoyo institucional, así como a Josep Tarrés su colaboración en el proceso de recogida de datos.
Los autores declaran que no se utilizaron herramientas de inteligencia artificial generativa en la redacción, el análisis de los datos ni la interpretación de este manuscrito, de acuerdo con las políticas de publicación de Apunts.
Financiación
Este estudio fue financiado por el Instituto Nacional de Educación Física de Cataluña (INEFC) de la Generalitat de Catalunya. La entidad financiadora no participó en el diseño del estudio ni en la recogida y el análisis de los datos ni en la decisión de publicar ni en la preparación del manuscrito.
Referencias
[1] Alt, T., Heinrich, K., Funken, J., & Potthast, W. (2015). Lower extremity kinematics of athletics curve sprinting. Journal of Sports Sciences, 33(6), 552–560. doi.org/10.1080/02640414.2014.960881
[2] Bell, D. R., Sanfilippo, J. L., Binkley, N., & Heiderscheit, B. C. (2014). Lean mass asymmetry influences force and power asymmetry during jumping in collegiate athletes. Journal of Strength and Conditioning Research, 28(4), 884–891. doi.org/10.1519/JSC.0000000000000367
[3] Bini, R., & Hume, P. (2015). Relationship between pedal force asymmetry and performance in cycling time trial. Journal of Sports Medicine and Physical Fitness, 55(9), 892–898.
[4] Bishop, C., Read, P., Brazier, J., Jarvis, P., Chavda, S., Bromley, T., & Turner, A. (2021). Effects of interlimb asymmetries on acceleration and change of direction speed: A between-sport comparison of professional soccer and cricket athletes. Journal of Strength and Conditioning Research, 35(8), 2095–2101. doi.org/10.1519/JSC.0000000000003135
[5] Bishop, C., Read, P., Bromley, T., Brazier, J., Jarvis, P., Chavda , S., & Turner, A. (2022). The association between interlimb asymmetry and athletic performance tasks: A season-long study in elite academy soccer players. Journal of Strength and Conditioning Research, 36(3), 787–795. doi.org/10.1519/JSC.0000000000003526
[6] Bishop, C., Turner, A., & Read, P. (2017). Effects of inter-limb asymmetries on physical and sports performance: A systematic review. Journal of Sports Sciences, 36(10), 1135–1144. doi.org/10.1080/02640414.2017.1361894
[7] Branski, L. K., Norbury, W. B., Herndon, D. N., Chinkes, D. L., Cochran, A., Suman, O., Benjamin, D., & Jeschke, M. G. (2010). Measurement of body composition in burned children: Is there a gold standard? JPEN: Journal of Parenteral and Enteral Nutrition, 34(1), 55–63. doi.org/10.1177/0148607109336601
[8] Cerezuela-Espejo, V., Hernández-Belmonte, A., Courel-Ibáñez, J., Conesa-Ros, E., Mora-Rodríguez, R., & Pallarés, J. G. (2021). Are we ready to measure running power? Repeatability and concurrent validity of five commercial technologies. European Journal of Sport Science, 21(3), 341–350. doi.org/10.1080/17461391.2020.1748117
[9] Chang, Y. H., & Kram, R. (2007). Limitations to maximum running speed on flat curves. Journal of Experimental Biology, 210(6), 971–982. doi.org/10.1242/jeb.02728
[10] Chilibeck, P. D., Paterson, D. H., Cunningham, D. A., Taylor, A. W., & Noble, E. G. (1997). Muscle capillarization, O2 diffusion distance, and VO₂ kinetics in old and young individuals. Journal of Applied Physiology, 82(1), 63–69. doi.org/10.1152/jappl.1997.82.1.63
[11] Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). Routledge. doi.org/10.4324/9780203771587
[12] Connick, M. J., & Li, F. X. (2015). Prolonged cycling alters stride time variability and kinematics of a post-cycle transition run in triathletes. Journal of Electromyography and Kinesiology, 25(1), 34–39. doi.org/10.1016/j.jelekin.2014.08.009
[13] D’Hondt, J., Chapelle, L., Bishop, C., Aerenhouts, D., Pauw, K.D., Clarys, P., & D’Hondt, E. (2024). Association between inter-limb asymmetry and endurance running performance in healthy populations: A systematic review. Sports Med - Open, 10(127). doi.org/10.1186/s40798-024-00790-w
[14] Feldmann, A., Schmitz, R., & Erlacher, D. (2019). Near-infrared spectroscopy-derived muscle oxygen saturation on a 0% to 100% scale: Reliability and validity of the Moxy Monitor. Journal of Biomedical Optics, 24(11), 115001. doi.org/10.1117/1.JBO.24.11.115001
[15] Fiori, L., Castiglia, S. F., Chini, G., Draicchio, F., Sacco, F., Serrao, M., Tatarelli, A., Varrecchia, T., & Ranavolo, A. (2024). The lower limb muscle co-activation map during human locomotion: From slow walking to running. Bioengineering, 11(3), 288. doi.org/10.3390/bioengineering11030288
[16] Fox, K. T., Pearson, L. T., & Hicks, K. M. (2023). The effect of lower inter-limb asymmetries on athletic performance: A systematic review and meta-analysis. PLoS One, 18(6), e0286942. doi.org/10.1371/journal.pone.0286942
[17] García, G. C., & Secchi, J. D. (2013). Relationship between the final speeds reached in the 20 metre Course Navette and the MAS-EVAL test. A proposal to predict the maximal aerobic speed. Apunts Medicina de l’Esport, 48(177), 27–34. doi.org/10.1016/j.apunts.2011.11.004
[18] Gilgen-Ammann, R., Taube, W., & Wyss, T. (2017). Gait asymmetry during 400- to 1000-m high-intensity track running in relation to injury history. International Journal of Sports Physiology and Performance, 12(S2), S2-157-S2-160. doi.org/10.1123/ijspp.2016-0379
[19] Grassi, B., Quaresima, V., Marconi, C., Ferrari, M., & Cerretelli, P. (1999). Blood lactate accumulation and muscle deoxygenation during incremental exercise. Journal of Applied Physiology, 87(1), 348–355. doi.org/10.1152/jappl.1999.87.1.348
[20] Hamill, J., Murphy, M., & Sussman, D. (1987). The effects of track turns on lower extremity function. International Journal of Sport Biomechanics, 3(3), 276–286. doi.org/10.1123/ijsb.3.3.276
[21] Heiden, T., & Burnett, A. (2003). The effect of cycling on muscle activation in the running leg of an Olympic distance triathlon. Sports Biomechanics, 2(1), 35–49. doi.org/10.1080/14763140308522806
[22] Helme, M., Tee, J., Emmonds, S., & Low, C. (2021). Does lower-limb asymmetry increase injury risk in sport? A systematic review. Physical Therapy in Sport, 49, 204–213. doi.org/10.1016/j.ptsp.2021.03.001
[23] Jacques, T., Bini, R., & Arndt, A. (2021). Running after cycling induces inter-limb differences in muscle activation but not in kinetics or kinematics. Journal of Sports Sciences, 39(2), 154–160. doi.org/10.1080/02640414.2020.1809176
[24] Karamanidis, K., Arampatzis, A., & Brüggemann, G.-P. (2003). Symmetry and reproducibility of kinematic parameters during various running techniques. Medicine and Science in Sports and Exercise, 35(6), 1009–1016. doi.org/10.1249/01.MSS.0000069337.49567.F0
[25] Knapik, J. J., Bauman, C. L., Jones, B. H., Harris, J. M., & Vaughan, L. (1991). Preseason strength and flexibility imbalances associated with athletic injuries in female collegiate athletes. American Journal of Sports Medicine, 19(1), 76–81. doi.org/10.1177/036354659101900113
[26] Léger, L., & Boucher, R. (1980). An indirect continuous running multistage field test: the Université de Montréal track test. Canadian journal of applied sport sciences. Journal canadien des sciences appliquées au sport, 5(2), 77–84.
[27] Loturco, I., Pereira, L. A., Kobal, R., Abad, C. C. C., Rosseti, M., Carpes, F. P., & Bishop, C. (2019). Do asymmetry scores influence speed and power performance in elite female soccer players? Biology of Sport, 36(3), 209–216. doi.org/10.5114/biolsport.2019.85454
[28] McManus, C. J., Collison, J., & Cooper, C. E. (2018). Performance comparison of the MOXY and PortaMon near-infrared spectroscopy muscle oximeters at rest and during exercise. Journal of Biomedical Optics, 23(1), 015007. doi.org/10.1117/1.JBO.23.1.015007
[29] Millet, G. P., & Vleck, V. E. (2000). Physiological and biomechanical adaptations to the cycle to run transition in Olympic triathlon: review and practical recommendations for training. British journal of sports medicine, 34(5), 384–390. doi.org/10.1136/bjsm.34.5.384
[30] Olcina, G., Perez-Sousa, M. Á., Escobar-Alvarez, J. A., & Timón, R. (2019). Effects of cycling on subsequent running performance, stride length, and muscle oxygen saturation in triathletes. Sports, 7(5), 115. doi.org/10.3390/sports7050115
[31] Orcioli-Silva, D., Beretta, V. S., Santos, P. C. R., Rasteiro, F. M., Marostegan, A. B., Vitório, R., Gobatto, C. A., & Manchado-Gobatto, F. B. (2024). Cerebral and muscle tissue oxygenation during exercise in healthy adults: A systematic review. Journal of sport and health science, 13(4), 459–471. doi.org/10.1016/j.jshs.2024.03.003
[32] Patoz, A., Blokker, T., Pedrani, N., Spicher, R., Borrani, F., & Malatesta, D. (2023). Biomechanical adaptations during exhaustive runs at 90 to 120% of peak aerobic speed. Scientific Reports, 13, 8236. doi.org/10.1038/s41598-023-35345-8
[33] Pauw, K.D., Roelands, B., Cheung, S. S., de Geus, B., Rietjens, G., & Meeusen, R. (2013). Guidelines to classify subject groups in sport-science research. International Journal of Sports Physiology and Performance, 8(2), 111–122. doi.org/10.1123/ijspp.8.2.111
[34] Richardson, J. T. (2011). Eta squared and partial eta squared as measures of effect size in educational research. Educational research review, 6(2), 135–147. doi.org/10.1016/j.edurev.2010.12.001
[35] Sendra-Pérez, C., Priego-Quesada, J. I., Murias, J. M., Carpes, F. P., Salvador-Palmer, R., & Encarnación-Martínez, A. (2025). Evaluation of leg symmetry in muscle oxygen saturation during submaximal to maximal cycling exercise. European Journal of Sport Science, 25(1), e12230. doi.org/10.1002/ejsc.12230
[36] Skotzke, P., Schwindling, S., & Meyer, T. (2024). Side differences and reproducibility of the Moxy muscle oximeter during cycling in trained men. European Journal of Applied Physiology, 124, 3075–3083. doi.org/10.1007/s00421-024-05514-2
[37] Soares, R. N., George, M. A., Proctor, D. N., & Murias, J. M. (2018). Differences in vascular function between trained and untrained limbs assessed by near-infrared spectroscopy. European Journal of Applied Physiology, 118, 2241–2248. doi.org/10.1007/s00421-018-3955-3
[38] Soto, A., Camerino, O., Iglesias, X., Anguera, M. T., & Castañer, M. (2019). LINCE PLUS: Research software for behaviour video analysis. Apunts. Educación Física y Deportes, 137, 149–153. doi.org/10.5672/apunts.2014-0983.es.(2019/3).137.11
[39] Tomczak, E. & Tomczak, M. (2014). The need to report effect size estimates revisited. An overview of some recommended measures of effect size. TRENDS in Sport Sciences, 21(1).
[40] van der Zwaard, S., Jaspers, R. T., Blokland, I. J., Achterberg, C., Visser, J. M., den Uil, A. R., Hofmijster, M. J., Levels, K., Noordhof, D. A., de Haan, A., de Koning, J. J., van der Laarse, W. J., & de Ruiter, C. J. (2016). Oxygenation Threshold Derived from Near-Infrared Spectroscopy: Reliability and Its Relationship with the First Ventilatory Threshold. PloS one, 11(9), e0162914. doi.org/10.1371/journal.pone.0162914
[41] van Rassel, C. R., Ajayi, O. O., Sales, K. M., Griffiths, J. K., Fletcher, J. R., Edwards, W. B., & MacInnis, M. J. (2023). Is running power a useful metric? Quantifying training intensity and aerobic fitness using Stryd running power near the maximal lactate steady state. Sensors, 23(21), 8729. doi.org/10.3390/s23218729
[42] Vasquez-Bonilla, A. A., Brazo-Sayavera, J., Timón, R., & Olcina, G. (2022). Monitoring muscle oxygen asymmetry as a strategy to prevent injuries in footballers. Research Quarterly for Exercise and Sport, 94(3), 609–617. doi.org/10.1080/02701367.2022.2026865
[43] Vasquez-Bonilla, A., Tomas-Carus, P., Brazo-Sayavera, J., Malta, J., Folgado, H., & Olcina, G. (2023). Muscle oxygenation is associated with bilateral strength asymmetry during isokinetic testing in sport teams. Science & Sports, 38(4), 426.e1–426.e9. doi.org/10.1016/j.scispo.2022.03.014
[44] World Athletics. (2019). Track and field facilities manual: Chapters 1–3. World Athletics. worldathletics.org/about-iaaf/documents/technical-information
[45] Yanci, J. (2014). Muscle strength and leg asymmetries in elite runners and cyclists: Original research article. International SportMed Journal, 15(3). hdl.handle.net/10520/EJC159075
ISSN: 2014-0983
Recibido: 16 de octubre de 2025
Aceptado: 27 de febrero de 2026
Publicado: 1 de julio de 2026
Editado por: © Generalitat de Catalunya Departament de la Presidència Institut Nacional d’Educació Física de Catalunya (INEFC)
© Copyright Generalitat de Catalunya (INEFC). Este artículo está disponible en la url https://www.revista-apunts.com/ Este trabajo está bajo la licencia Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. Las imágenes u otro material de terceros en este artículo se incluyen en la licencia Creative Commons del artículo, a menos que se indique lo contrario en la línea de crédito. Si el material no está incluido en la licencia Creative Commons, los usuarios deberán obtener el permiso del titular de la licencia para reproducir el material. Para ver una copia de esta licencia, visite https://creativecommons.org/licenses/by-nc-nd/4.0/deed.es_ES


