Activación de las fibras musculares de las extremidades inferiores en dos modalidades de baile latino
Encarnación Liébana
Cristina Monleón García
Carlos Barrios
Consuelo Moratal
*Correspondencia: Encarnación Liébana Giménez encarnacion.liebana@ucv.es
Citación
Liébana, E., Monleón, C., Barrios, C. & Moratal, C. (2024). Lower extremity muscle fibers activation in two Latin dance modalities. Apunts Educación Física y Deportes, 156, 57-65. https://doi.org/10.5672/apunts.2014-0983.es.(2024/2).156.07
Resumen
El objetivo principal de este estudio era investigar la actividad muscular de diferentes tipos de fibras musculares entre los estilos de baile rumba y jive en dieciocho deportistas de baile deportivo de élite (edad media: 19.6 ± 3.2 años). Las mediciones se realizaron mediante electromiografía de superficie (EMG) durante la ejecución de la coreografía. Se registró y analizó la EMG en ambas piernas del recto femoral (RF), el bíceps femoral (BF), el tibial anterior (TA) y el gastrocnemio medial (GM). En la rumba, la activación total del RF (mediana, 115.95; IQR, 36.00 mV) fue menor que la del BF (mediana, 146.68; IQR, 10.02 mV; p = .002) y el GM (mediana, 149.81; IQR, 85.66 mV; p = .035). En el jive, la mayor activación global correspondió al BF (mediana, 155.40; IQR, 44.89 mV), y las diferencias tuvieron significación estadística en comparación con la activación del TA (mediana, 123.09; IQR, 51.24 mV; p = .028). Se encontraron diferencias significativas entre la rumba y el jive en las fibras tipo I del RF (p ≤ .05), las fibras tipo IIa del TA (p ≤ .05); y las fibras tipo IIb del GM (p ≤ .05), tanto en hombres como en mujeres. Entre los bailarines, hubo diferencias en las fibras de tipo IIb del GM (p ≤ .05), mientras que se observaron diferencias en las fibras tipo I del TA entre las mujeres (p ≤ .05). Este estudio muestra evidencias experimentales de una activación muscular significativamente distinta para la extremidad inferior en bailes con diferente tempo. Los resultados de este estudio aportan información relevante para optimizar los programas de entrenamiento de alto rendimiento y prevención de lesiones, que es clave para el éxito profesional en el baile deportivo.
Introducción
El baile deportivo es una combinación de arte, deporte y rendimiento deportivo. Se realiza por parejas, por lo que pueden expresar emociones y formar movimientos armoniosos en respuesta a diferentes tipos de música (Lukić et al., 2011; Riding et al., 2013; Uzunović et al., 2009; Uzunović y Kostić, 2005).
Este deporte se compone de tres especialidades: bailes latinos, estándares y los “10 bailes”; estos últimos son una combinación de bailes estándares y latinos (WDSF, 2017). Los bailes latinos se caracterizan principalmente por figuras abiertas, o semiabiertas, y cerradas que requieren una conexión visual (Čačković et al., 2012). Los bailes que conforman la especialidad latina son: la samba, con un tempo de 50–52 pulsaciones por minuto (p.p.m.); el chachachá, con un tempo de 30–32 p.p.m.; la rumba, con un tempo de 25–27 p.p.m.; el pasodoble, con un tempo de 60–62 p.p.m.; y el jive, con un tempo de 42–44 p.p.m. (FEBD, 2017).
La rumba es un baile con ritmo corporal muy expresivo en el que se representan movimientos atractivos y delicados. Destaca la importancia de la fluidez y el ritmo del movimiento en los músculos de la espalda, junto con los movimientos de cadera de los bailarines (Shang, 2013). De este modo, las caderas dibujan de forma natural una trayectoria de movimiento en un “8” invertido controlado por los bailarines (Shang, 2013).
El jive es el baile latino más explosivo y provoca una alta frecuencia cardíaca (Bria et al., 2011; Liiv et al., 2014). Fundamentalmente, el jive se caracteriza por el equilibrio entre el SWINGy (con giro) y el JUMPy (con salto), dos principios básicos que contribuyen al rendimiento general del baile (Dance Comp Review, 2014).
El SWINGy está presente en cada uno de los pasos realizados en el jive. El cuerpo se inclina hacia delante, provocando un desplazamiento lateral mediante el movimiento de la cadera, y se mantiene erguido durante los pasos hacia delante o hacia atrás. El apoyo del pie se consigue principalmente mediante el apoyo del primer y segundo metatarsianos (Dance Comp Review, 2014).
En cuanto al JUMPy, la acción principal es la flexión de la rodilla simultánea a la contracción del abdomen, que da lugar a un pequeño salto. Se compone de cuatro fases: paso, salto, vuelo y aterrizaje. En la primera fase, el paso se realiza con una pequeña flexión en la rodilla, lo que da lugar a la siguiente fase: la extensión de la rodilla, que produce el impulso para saltar. Esto se traslada a la fase de vuelo y posteriormente a la de aterrizaje, donde la mayor parte del apoyo lo proporcionan los primeros metatarsianos con una ligera flexión de las rodillas para absorber el impacto (Dance Comp Review, 2014).
La electromiografía de superficie es una técnica habitual y no invasiva para analizar las contracciones musculares en aplicaciones reales (Hermens y Freriks, 1997; Liu et al., 2002). En los seres humanos, las fibras rápidas y lentas no están separadas físicamente, pero la evolución ha mantenido de algún modo la separación de los distintos tipos de fibras. Las unidades motoras del músculo humano se suelen clasificar en tres grupos diferentes comúnmente denominados: oxidativo lento o tipo I (TI); oxidativo rápido o tipo IIa (TIIa); y glucolítico rápido o tipo IIb (TIIb)(Von Tscharner y Goepfert, 2006; Brooke y Kaiser, 1970). Estos grupos pueden formarse en diferentes proporciones para distintos periodos de un movimiento y pueden explicar, al menos, una parte de la variabilidad espectral (Von Tscharner y Goepfert, 2003; Wakeling et al., 2001). Para analizar los datos obtenidos con la electromiografía de superficie en contracciones musculares variables, se han adoptado técnicas de medición de la frecuencia temporal (Kumar et al., 2003). La transformada de Wavelet continua (comparación de diferentes técnicas de frecuencia a lo largo del tiempo) produce resultados precisos con una buena representación de la localización temporal y frecuencial (Karlsson et al., 2000).
Existen escasas pruebas sobre el baile deportivo y, en particular, sobre la actividad muscular de los bailarines. Zagorc et al. (2010) utilizaron la tensiomiografía para estudiar el tiempo de contracción de deportistas de baile deportivo y observaron que el tiempo de contracción en músculos como el gastrocnemio variaba entre géneros. Liébana et al. (2017) analizaron los EMG de deportistas de baile deportivo que interpretaban la rumba bolero y observaron las diferencias. De este modo, se encontraron diferencias de activación en el recto femoral (RF) y el gastrocnemio medial (GM) de las mujeres asociadas con la rumba bolero, así como en el tibial anterior (TA) y el GM, además de diferencias entre los ritmos de varios géneros de baile (Haeufle et al., 2010).
Para comprender cómo el complejo sistema musculoesquelético puede generar una fuerza adecuada en las piernas, es necesario conocer las propiedades intrínsecas de los músculos (Haeufle et al., 2010). Por ello, la finalidad de nuestro estudio era evaluar la actividad muscular de los tres tipos de fibras musculares del RF, el BF, el TA y el GM de los bailarines, comparando la rumba con el jive en hombres y mujeres con el fin de diseñar entrenamientos específicos para estos deportistas. Los resultados previstos indicarían diferencias significativas en la activación de los tres tipos de fibras musculares analizadas: RF, BF, TA y GM (en ambas piernas, dominante y no dominante) en las extremidades inferiores de los bailarines, comparando la rumba con el jive, y entre géneros.
Materiales y metodología
Participantes
Los participantes fueron 18 deportistas de baile deportivo (9 parejas). Todos ellos son bailarines de categoría A (la máxima categoría) con 10.44 ± 3.51 años de experiencia en el baile; además, son especialistas en los “10 bailes” o en bailes latinos (véase la Tabla 2).
Los criterios de inclusión exigían que los participantes estuvieran en activo durante el estudio, tuvieran más de 18 años y se ajustaran a los 10 bailes o a la modalidad latina; los participantes no debían haber sufrido lesiones durante el año anterior y tenían que haber bailado en la categoría A durante al menos un año con la misma pareja. Se contactó con seis escuelas de danza que podían estar interesadas en participar en este estudio. De un total de 10 parejas que cumplían los criterios de inclusión, se midieron 9, es decir, 18 participantes, todos ellos con dominancia derecha. Una pareja fue excluida de las mediciones por problemas de agenda; además, estos bailarines disolvieron posteriormente su pareja.
Procedimientos
Se pidió a los bailarines que no realizaran ningún ejercicio físico en las 24 horas previas a la sesión del estudio. Durante la sesión, se tomaron medidas de la estatura (con un tallímetro SECA 709 7021994; Seca GmbH y Co. KG., Alemania) y datos antropométricos (peso e índice de masa corporal) mediante impedancia bioeléctrica (analizador de composición corporal segmentaria Tanita BC-418 MA; Tanita Corporation, Japón).
Para recopilar los datos con electromiografía (EMG), se siguió el protocolo estándar (véase la Tabla 1) para preparar la piel de los participantes y colocar los electrodos (Torrence y Compo, 1998; Welch, 1967).
Posteriormente, los participantes realizaron una adaptación idéntica al calentamiento RAMP, que consistió en integrar la movilidad articular en tobillos, rodillas, caderas y hombros, seguida de movimientos específicos por parejas sin música, y continuaron la activación mediante sentadillas y planchas; finalmente, las parejas realizaron un baile al ritmo de la música (Jeffreys, 2007). Se efectuaron mediciones mediante electromiografía de superficie de la activación muscular durante todas las coreografías competitivas (120 s cada tipo de baile), preparadas e interpretadas por los bailarines (Mega Electronics Ltd., Kuopio, Finlandia).
Existen trabajos previos con coreografías preestablecidas (Liébana et al., 2018), por lo que este estudio pretende medir la activación en un contexto ecológico, acercándonos al contexto real de competición.
El estudio fue aprobado por el Comité de Ética de la Investigación de la Universidad Católica de Valencia San Vicente Mártir, con el código UCV/2015-2016/60, y se ajusta a la Declaración de Helsinki. Los participantes conocían la finalidad del estudio y todos recibieron un consentimiento informado por escrito.

Tratamiento de datos
Todas las mediciones de EMG fueron recogidas por sensores Mega WBA con una frecuencia de muestreo de 1,000 Hz, un paso de banda de frecuencia del sensor entre 20 y 500 Hz, 200 electrodos de espuma Kendall con hidrogel adhesivo conductor (colocados con una distancia máxima entre electrodos de 20 mm) y se compilaron utilizando el software Megawin 3.1 (Mega Electronics Ltd., Kuopio, Finlandia). A continuación, se transfirieron a un archivo ASCII para su posterior análisis. Los datos extraídos de la activación muscular se expresan en milivoltios (mV). El archivo se transformó a .m para su análisis mediante Matlab.
El tratamiento de los datos se inició utilizando Matlab R2017b, que selecciona automáticamente los segundos centrales de cada ejercicio. La señal se filtró mediante el uso de un filtro pasabanda para establecer los valores mínimos con un límite de 20 Hz y valores máximos de 400 Hz. Se obtuvo la raíz cuadrática media (RMS). Se realizó una transformada de Fourier utilizando la transformada rápida de Fourier (Welch, 1967), que indica el espectro de potencia media y permite estimar la densidad espectral. Para ello, se empleó el periodograma de Welch con una ventana de Hamming de longitud 1012, a fin de estimar la densidad espectral (Welch, 1967). En este método, se lleva a cabo la fragmentación de la serie temporal, que permite calcular un periodograma modificado para cada uno de los segmentos. Una vez calculada la media, este proceso facilita la estimación de la densidad espectral. El método Welch es una mejora del método estándar del periodograma, ya que realiza una reducción del ruido en el espectro de potencia estimado. Sin embargo, este método plantea un problema. Para corregirlo, se aplicó un análisis de tiempo-frecuencia, en el que una ventana de longitud fija se desplaza a lo largo de la señal para relacionar las frecuencias con el tiempo y pueden evaluarse estas frecuencias en cada ventana.
A continuación, se aplica un análisis de series temporales no estacionarias mediante la transformada de Wavelet (Torrence y Compo, 1998), un método que permite analizar la señal en el dominio de la escala temporal. Se trata de una serie temporal formada por familias de funciones definidas temporal y espacialmente, que se producen por escalado y traslación de una función denominada “función base”. El escalograma consiste en un espectro de potencia promediado para las diferentes frecuencias o escalas, concedidas en cada valor temporal (Torrence y Compo, 1998).
La transformada de Wavelet se divide en dos variables: la transformada de Wavelet continua detecta patrones o modificaciones a lo largo de la evolución temporal de la señal a distintas escalas, mientras que la transformada de Wavelet discreta se obtiene mediante la descomposición de la señal en distintas zonas del espectro de frecuencias, seguida del filtrado de datos para obtener los coeficientes de Wavelet. El filtrado se produce en relación con la aproximación, el detalle y los filtros de paso bajo (5 Hz) y paso alto (250 Hz). Estos resultados son la descomposición de la señal global en señales ortogonales que permiten dividir las señales en cada una de las bandas de frecuencia. En este caso, se fijaron tres bandas: la primera, < 70 Hz; la segunda, entre 70 y 125 Hz; y la última, entre 126 y 250 Hz (Torrence y Compo, 1998).
Análisis estadístico
Para analizar los datos, se utilizó el paquete estadístico SPSS 22.0 (IBM, Chicago, Illinois, EE. UU.). Las características descriptivas de la antropometría se presentan como medias y desviaciones típicas (DT). Debido al tamaño limitado de la muestra (9 parejas), se recomendaron pruebas no paramétricas para comparar las variables cuantitativas. Los cambios en la activación muscular entre las dos modalidades de danza se evaluaron con la prueba de rangos de Wilcoxon. Dada la posible variabilidad de las mediciones de EMG en los participantes, los valores se presentaron como medianas y rangos intercuartílicos (IQR). Las comparaciones de variables cuantitativas entre bailarines y bailarinas se evaluaron con la prueba de Mann-Whitney. También se indicó el valor Z. Para todos los efectos principales e interacciones, se adoptó un nivel de confianza de .05.
Resultados
Las características antropométricas descriptivas de la muestra se presentan en la Tabla 2. No hubo diferencias significativas entre bailarines y bailarinas en las variables antropométricas (Tabla 2).
Considerando la activación en el EMG de todas las fibras musculares juntas en los diferentes músculos analizados, la rumba mostró menor activación que el jive en los músculos de la parte proximal de la extremidad inferior (RF y BF), aunque no hubo diferencias entre las dos modalidades de baile (Figura 1A). En la rumba, la activación del RF (mediana, 115.95; IQR, 36.00 mV) fue menor que la del BF (mediana, 146.68; IQR, 10.02 mV; p = .002) y el GM (mediana, 149.81; IQR, 85.66 mV; p = .035). El GM mostró la mayor activación muscular en la rumba. En el jive, la mayor activación correspondió al BF (mediana, 155.40; IQR, 44.89 mV), y las diferencias tuvieron significación estadística en comparación con la activación del TA (mediana, 123.09; IQR, 51.24 mV; p = .028). No hubo diferencias entre la pierna dominante y la no dominante en la activación en el EMG global de los músculos durante las dos modalidades de danza.
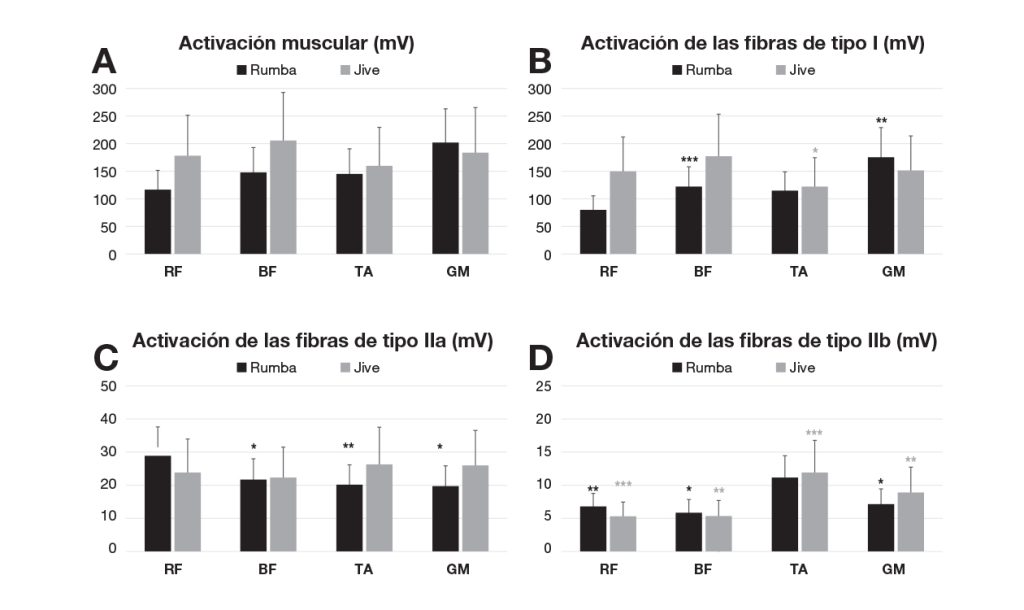
Nota: *p < .05; **p < .01; ***p = .001. A: registro mediante EMG de la activación global de los diferentes músculos analizados en la pierna dominante; B: activación de las fibras de tipo I; C: activación de las fibras de tipo IIa; y D: activación de las fibras de tipo IIb en la pierna dominante. (RF: Recto femoral; BF: Bíceps femoral; TA: Tibial anterior; GM: Gastrocnemio medial).
En la figura 1, B, C y D muestran una comparación más detallada entre la activación de diferentes tipos de fibras en los músculos evaluados en la pierna dominante en relación con la modalidad de danza. En la rumba, la activación de las fibras de tipo I del RF fue significativamente menor que en el BF (z = -3.201; p = .001) y GM (z = -2.635; p = .008). Además, el GM mostró la mayor activación muscular de fibras de tipo I en la rumba. En el jive, la mayor activación correspondió al BF (mediana, 124.05; IQR, 55.35 mV), y las diferencias tuvieron significación estadística en comparación con la activación del TA (mediana, 85.90; IQR, 35.52 mV; p = .028).
En cuanto a las fibras de tipo IIa, el músculo RF mostró una mayor activación en la rumba que en el jive. En la rumba, la activación del RF fue significativamente mayor que la del BF (z = -2.3301; p = .020), el TA (z = -2.809; p = .005) y el GM (z = -2.243; p = .025) (Fig. 1C).
La activación de las fibras de tipo IIb fue muy baja en todos los músculos estudiados. En ambas modalidades de danza, la mayor activación se encontró en el TA (Figura 1D). En la rumba, la activación del TA mostró diferencias con significación estadística en comparación con el RF (z = -2.940; p = .003), el BF (z = -2.461; p = .014) y el GM (z = -2.025; p = .043). En el jive, la activación del TA también mostró diferencias con significación estadística en comparación con el RF (z = -3.201; p = .001) y el BF (z = -3.157; p = .002). En el jive, también hubo diferencias entre la activación del RF y del GM (z = -2.765; p = .006), y entre el BF y el GM (z = -3.157 p = .002).
En la Tabla 3, se presentan los datos descriptivos sobre la activación de los diferentes tipos de fibras musculares, tanto en la pierna dominante como en la no dominante, durante las dos modalidades de danza. En cuanto a las fibras musculares de tipo I, la rumba mostró menor activación que el jive en los músculos RF y BF de la pierna dominante, pero las diferencias solo fueron significativas para el RF (p = .022). En las fibras de tipo IIa, el TA y el GM de la pierna dominante mostraron mayor activación en el jive que en la rumba, pero las diferencias solo tuvieron significación estadística en el TA (p = .002). La activación de las fibras de tipo IIb fue muy baja en todos los músculos estudiados. Las diferencias entre la rumba y el jive solo se detectaron en el GM de la pierna no dominante (p = .016). Cuando se compararon las piernas dominantes y no dominantes dentro de cada modalidad de baile, solo hubo diferencias en la activación de las fibras de tipo I del GM, que fueron mayores en la pierna dominante en la rumba (p = .006).
En el jive, no hubo diferencias en la activación de los distintos músculos analizados. Los músculos distales (TA y GM) se activaron más en el jive que en la rumba. En el jive, la mayor activación de fibras de tipo IIa se detectó en el GM en ambos lados (Tabla 3).
En la Tabla 4, se muestran los datos descriptivos relativos a los tipos de fibras para la rumba y el jive en ambos sexos. Se obtuvieron diferencias significativas para hombres y mujeres en las fibras tipo I del TA, tanto en rumba como en jive, en la pierna dominante (p = .028). En las mujeres, hubo diferencias significativas en la activación no dominante del TA en la rumba y el jive (p = .038). Cuando se compararon bailarines y bailarinas dentro de cada modalidad, solo se encontraron diferencias en la activación EMG en el GM de la pierna dominante durante la rumba (Tabla 4). Los hombres mostraron una menor activación de las fibras de tipo I en el GM (p = .019) y una mayor activación de las fibras de tipo IIa en el GM (p = .014) que las mujeres.
Discusión
Los bailarines en general, y particularmente el baile deportivo, no se han estudiado en profundidad y existe una notable falta de información sobre el comportamiento de las fibras musculares durante la práctica de este deporte. La importancia del presente estudio radica en las diferencias significativas identificadas entre la rumba y el jive, las cuales facilitan la programación y planificación del entrenamiento neuromuscular según nuestros resultados, si bien somos conscientes de las limitaciones debidas al reducido número de la muestra, pues se trata de un deporte minoritario. Estos bailes presentan diferencias importantes en cuanto a técnica, ritmo y tempo, pero el hecho de que existan claras diferencias en la activación muscular también debe tenerse en cuenta en el desarrollo de las sesiones de entrenamiento para optimizar el rendimiento y reducir el riesgo de lesiones.
La técnica, el ritmo y los movimientos de la rumba son más lentos que en el jive. En la rumba, se busca una estética particular, por lo que su técnica favorece la extensión de las extremidades inferiores (Shang, 2013). En cambio, el jive tiene características similares a los saltos a la pata coja, con sus patadas y saltos consecutivos. Este ejercicio combina velocidad y fuerza para producir un movimiento explosivo-reactivo (Cappa y Behm, 2013). Estos ejercicios implican contracciones musculares excéntricas (estiramiento) y concéntricas (acortamiento), generalmente utilizando el cuerpo como sobrecarga y generando un ciclo de estiramiento-acortamiento (SSC) (Cappa y Behm, 2013). Para que una acción muscular se clasifique como SSC durante la actividad de rebote, el patrón de activación muscular debe incluir una preactivación previa al contacto con el suelo, una acción excéntrica rápida y una transición inmediata y rápida entre las fases excéntrica y concéntrica (Komi, 2000). Por esta razón, existen diferencias significativas entre la rumba y el jive en RF1_TI, TA1_TIIa y GM2_TIIb en la pierna no dominante. Al igual que en los saltos a la pata coja, en el jive los músculos isquiotibiales y los cuádriceps deben estar activos al mismo tiempo para crear estabilidad (Wibawa et al., 2016).
El TA es un músculo que se activa no solo en la dorsiflexión del pie, sino también para controlar la pronación, por lo que en el jive estaría actuando excéntricamente para contribuir a ambos movimientos (Cappa y Behm, 2013). Por ende, las diferencias entre la rumba y el jive se encuentran principalmente en el TA1_TIIa. Las fibras del TA1_TIIa se activan en movimientos cortos y de alta intensidad, ya que este músculo proporciona apoyo mediante la activación excéntrica de frenado. Esta activación también se ve favorecida para el apoyo y cuando los bailarines realizan una técnica de aterrizaje utilizando el primer y segundo metatarsianos (Dance Comp Review, 2014).
También se encontraron diferencias significativas entre la rumba y el jive en relación con el GM2_TIIb. Esto puede deberse al trabajo de SSC de este músculo, ya que una técnica de jive favorece la realización continua de saltos rápidos y patadas, generando así trabajo pliométrico y la realización de un SSC (Cappa y Behm, 2013). Nicol et al. (2006) destacan en su trabajo que el gastrocnemio reacciona de forma diferente en el aterrizaje tras un salto; si el salto es pequeño, las fibras del gastrocnemio tienden a acortarse para frenar. Si el salto es alto y requiere un frenado importante, las fibras musculares tienden a alargarse. Esto se debe a la menor resistencia al estiramiento por la posible liberación de puentes transversales (Nicol et al., 2006). La carga de impacto determina el comportamiento del fascículo en un músculo específico y la intensidad del esfuerzo tras la fase de frenado tiene cierta influencia en esta interacción al afectar al retroceso del tendón en el empuje final (Nicol et al., 2006).
Por otra parte, nuestros resultados muestran diferencias en la activación muscular entre hombres y mujeres. Estos datos estarían en consonancia con los obtenidos en el estudio de Liébana et al. (2017). Se observaron discrepancias en la activación muscular entre sexos en el tibial anterior y el gastrocnemio, lo cual demuestra que la activación muscular entre hombres y mujeres es diferente (Haeufle et al., 2010). Esto puede deberse tanto a las diferencias entre los pasos de las mujeres y los de los hombres como a la marcada diferencia entre los tacones de los zapatos de baile de hombres y mujeres que, en consecuencia, provoca que los músculos generen fuerzas activas diferentes en las piernas. Hill (1938) describió y distinguió las propiedades intrínsecas de un solo músculo, representado por un elemento elástico en serie y un elemento contráctil con relaciones fuerza-longitud y fuerza-velocidad. Las propiedades musculares pueden compensar las perturbaciones y facilitar la convergencia de los movimientos dinámicos y explosivos. Las propiedades intrínsecas del músculo representadas por la función fuerza-longitud-velocidad en los modelos musculares de Hill actúan como un sistema de retroalimentación periférica sin retardo (Haeufle et al., 2010).
Las diferencias en la activación muscular entre los tipos de fibras, músculos, sexos y bailes observados en nuestro estudio demostrarían la necesidad de un entrenamiento individualizado y planificado para cada tipo de baile y pareja (hombre frente a mujer). De ahí la necesidad de un entrenamiento específico para bailarines que aborde el tipo de fuerza trabajada, el entrenamiento de la movilidad, la técnica y el control motor. Estos factores son de gran importancia para prevenir lesiones y lograr el máximo rendimiento deportivo.
En relación con los datos mostrados en este estudio, cabe destacar las limitaciones en cuanto a la muestra, ya que se trata de un número bajo de participantes en comparación con otros estudios, pero representativo del número de participantes en el campeonato de España, teniendo en cuenta que el baile deportivo es un deporte minoritario.
Conclusiones
Los resultados de este estudio proporcionan datos fácticos experimentales de activaciones musculares significativamente diferentes para la extremidad inferior en función del sexo y la modalidad de danza. Se encontraron diferencias significativas en la activación en función del tipo de fibra entre rumba y jive. Desde una perspectiva clínica, nuestros resultados pueden ayudar a entrenadores y médicos deportivos a comprender el perfil deportivo específico de los bailarines de élite. El conocimiento de la función muscular, la actividad y el equilibrio es crucial para optimizar el alto nivel de rendimiento de estos deportistas, así como para favorecer los programas de prevención de lesiones, que son cruciales para maximizar su éxito deportivo. Estos resultados suponen un primer paso para proporcionar valores de referencia de las fibras musculares implicadas en los movimientos de los deportes de danza; dichos valores pueden contribuir al diseño de ejercicios que favorezcan el rendimiento deportivo y la prevención de lesiones. Al realizar este tipo de análisis mediante transformadas de Wavelet, se ha podido observar que este análisis es sensible y muestra resultados coherentes, teniendo en cuenta en todo momento las limitaciones del estudio. Pueden utilizarse transformadas de Wavelet para el análisis de movimientos con características similares (Cappa y Behm, 2013). Este análisis permite comprender las exigencias musculares de los distintos tipos de danza. Por ende, para el correcto desarrollo del jive es importante el trabajo pliométrico, con un ciclo de acortamiento-estiramiento. Asimismo, para minimizar el desequilibrio muscular entre la cadena posterior y la anterior, así como la asimetría entre la pierna dominante y la no dominante, se recomendaría el trabajo de fuerza.
Referencias
[1] Bria, S., Bianco, M., Galvani, C., Palmieri, V., Zeppilli, P., & Faina, M. (2011). Physiological characteristics of elite sport-dancers. The Journal of Sports Medicine and Physical Fitness, 51(2), 194-203.
[2] Brooke, M. H., & Kaiser, K. K. (1970). Muscle Fiber Types: How Many and What Kind? Archives of Neurology, 23(4), 369-379. doi.org/10.1001/archneur.1970.00480280083010
[3] Čačković, L., Baric, R., & Vlasic, J. (2012). Psychological stress in dancesport. Acta Kinesiologica, 6(2), 71-74. actakin.com/PDFS/BR0602/SVEE/04%20CL%2013%20LC.pdf
[4] Cappa, D. F., & Behm, D. G. (2013). Neuromuscular Characteristics of Drop and Hurdle Jumps With Different Types of Landings: Journal of Strength and Conditioning Research, 27(11), 3011-3020. doi.org/10.1519/JSC.0b013e31828c28b3
[5] Criswell, E., & Cram, J. R. (Eds.). (2011). Cram’s introduction to surface electromyography (2nd ed). Jones and Bartlett.
[6] Dance Comp Review. (2014). Principals of International Latin Jive. Dance Comp Review. dancecompreview.com/principals-international-latin-jive/
[7] FEBD. (2017). Código Básico de Competiciones. www.febd.es/wordpress/wp-content/uploads/2017/01/codigo_basico_de_competiciones_11_5.pdf
[8] Haeufle, D. F. B., Grimmer, S., & Seyfarth, A. (2010). The role of intrinsic muscle properties for stable hopping—Stability is achieved by the force–velocity relation. Bioinspiration & Biomimetics, 5(1), 016004. doi.org/10.1088/1748-3182/5/1/016004
[9] Hermens, H., & Freriks, B. (1997). The state of the art on sensors and sensor placement procedures for surface electromyography: A proposal for sensor placement procedures. Roessingh Research and Development.
[10] Hill, A. V. (1938). The Heat of Shortening and the Dynamic Constants of Muscle. Proceedings of the Royal Society B: Biological Sciences, 126(843), 136-195. doi.org/10.1098/rspb.1938.0050
[11] Jeffreys, I. (2007). Warm up revisited – the ‘ramp’ method of optimising performance preparation. Professional Strength and Conditioning, 6, 12-18.
[12] Karlsson, S., Jun Yu, & Akay, M. (2000). Time-frequency analysis of myoelectric signals during dynamic contractions: A comparative study. IEEE Transactions on Biomedical Engineering, 47(2), 228-238. doi.org/10.1109/10.821766
[13] Komi, P. V. (2000). Stretch-shortening cycle: A powerful model to study normal and fatigued muscle. Journal of Biomechanics, 33(10), 1197-1206. doi.org/10.1016/S0021-9290(00)00064-6
[14] Kumar, D. K., Pah, N. D., & Bradley, A. (2003). Wavelet analysis of surface electromyography. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 11(4), 400-406. doi.org/10.1109/TNSRE.2003.819901
[15] Liébana, E., Blasco, E., Monleón, C., Pablos, C., & Moratal, C. (2017). Muscular activation in rumba bolero in elite dancers of DanceSport. Journal of Human Sport & Exercise, 12(3proc), S807-S812. doi.org/10.14198/jhse.2017.12.Proc3.04
[16] Liébana, E., Monleón, C., Morales, R., Pablos, C., Moratal, C., & Blasco, E. (2018). Muscle Activation in the Main Muscle Groups of the Lower Limbs in High-Level Dancesport Athletes. Medical Problems of Performing Artists, 33(4), 231-237. doi.org/10.21091/mppa.2018.4034
[17] Liiv, Jürimäe, T., Mäestu, J., Purge, P., Hannus, A., & Jürimäe, J. (2014). Physiological characteristics of elite dancers of different dance styles. European Journal of Sport Science, 14, S429-S436. doi.org/10.1080/17461391.2012.711861
[18] Liu, J. Z., Brown, R. W., & Yue, G. H. (2002). A dynamical model of muscle activation, fatigue, and recovery. Biophysical Journal, 82(5), 2344-2359. www.ncbi.nlm.nih.gov/pmc/articles/PMC1302027/
[19] Lukić, A., Bijelić, S., Zagorc, M., & Zuhrić-Sebić, L. (2011). The importance of strength in sport dance performance technique. Sportlogia, 7(1), 61-67. doi.org/10.5550/sgia.110701.en.061L
[20] Nicol, C., Avela, J., & Komi, P. V. (2006). The Stretch-Shortening Cycle. Sports Medicine, 36(11), 977-999. doi.org/10.2165/00007256-200636110-00004
[21] Riding, T., Wyon, M., Ambegaonkar, J., & Redding, E. (2013). A Bibliographic Review of Medicine and Science Research in DanceSport. Medical Problems of Performing Artists, 28(2), 70-79. doi.org/10.21091/mppa.2013.2013
[22] Shang, Y. (2013). Technical analysis of the hips squeezing action in rumba based on biomechanics. Trade Science Inc, 8(9), 1205-1209.
[23] Torrence, C., & Compo, G. P. (1998). A practical guide to wavelet analysis. Bulletin of the American Meteorological Society, 79(1), 61-78. journals.ametsoc.org/doi/abs/10.1175/1520-0477(1998)079%3C0061:APGTWA%3E2.0.CO;2
[24] Uzunović, S., & Kostić, R. (2005). A study of success in A study of success in Latin American sport dancing. Facta Universitatis, Series Physical Education and Sport 3, 23-35.
[25] Uzunović, S., Kostić, R., & Miletić, Đ. (2009). Motor status of competitive young sport dancers-gender differences. Acta Kinesiologica, 3(1), 83-88. www.actakin.com/PDFS/BR0301/SVEE/04%20CL%2014%20SU.pdf
[26] Von Tscharner, V., & Goepfert, B. (2003). Gender dependent EMGs of runners resolved by time/frequency and principal pattern analysis. Journal of Electromyography and Kinesiology, 13(3), 253-272. doi.org/10.1016/S1050-6411(02)00111-6
[27] Von Tscharner, V., & Goepfert, B. (2006). Estimation of the interplay between groups of fast and slow muscle fibers of the tibialis anterior and gastrocnemius muscle while running. Journal of Electromyography and Kinesiology, 16(2), 188-197. doi.org/10.1016/j.jelekin.2005.07.004
[28] Wakeling, J. M., Pascual, S. A., Nigg, B. M., & Tscharner, V. (2001). Surface EMG shows distinct populations of muscle activity when measured during sustained sub-maximal exercise. European Journal of Applied Physiology, 86(1), 40-47. doi.org/10.1007/s004210100508
[29] WDSF. (2017). World Dance Sport Federation. About DanceSport. www.worlddancesport.org/about
[30] Welch, P. (1967). The use of fast Fourier Transform for the Estimation of Power Spectra: A Method based on time avering over short, modilied periodograms. 15, 70-73. doi.org/10.1109/TAU.1967.1161901
[31] Wibawa, A. D., Verdonschot, N., Halbertsma, J. P. K., Burgerhof, J. G. M., Diercks, R. L., & Verkerke, G. J. (2016). Musculoskeletal modeling of human lower limb during normal walking, one-legged forward hopping and side jumping: Comparison of measured EMG and predicted muscle activity patterns. Journal of Biomechanics, 49(15), 3660-3666. doi.org/10.1016/j.jbiomech.2016.09.041
[32] Zagorc, M., Šimunič, B., Pišot, R., & Oreb, G. (2010). A comparison of contractile parameters among twelve skeletal muscles of inter-dance couples. Kinesiologia Slovenica, 16(3), 57-65.
ISSN: 2014-0983
Recibido: 2 de junio de 2023
Aceptado: 10 de octubre de 2023
Publicado: 1 de abril de 2024
Editado por: © Generalitat de Catalunya Departament de la Presidència Institut Nacional d’Educació Física de Catalunya (INEFC)
© Copyright Generalitat de Catalunya (INEFC). Este artículo está disponible en la url https://www.revista-apunts.com/ Este trabajo está bajo la licencia Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. Las imágenes u otro material de terceros en este artículo se incluyen en la licencia Creative Commons del artículo, a menos que se indique lo contrario en la línea de crédito. Si el material no está incluido en la licencia Creative Commons, los usuarios deberán obtener el permiso del titular de la licencia para reproducir el material. Para ver una copia de esta licencia, visite https://creativecommons.org/licenses/by-nc-nd/4.0/deed.es_ES


