Activació de les fibres musculars de les extremitats inferiors en dues modalitats de ball llatí
Encarnación Liébana
Cristina Monleón García
Carlos Barrios
Consuelo Moratal
*Correspondència: Encarnación Liébana Giménez encarnacion.liebana@ucv.es
Citació
Liébana, E., Monleón, C., Barrios, C. & Moratal, C. (2024). Lower extremity muscle fibers activation in two Latin dance modalities. Apunts Educación Física y Deportes, 156, 57-65. https://doi.org/10.5672/apunts.2014-0983.es.(2024/2).156.07
Resum
L’objectiu principal d’aquest estudi era investigar l’activitat muscular de diferents tipus de fibres musculars entre els estils de ball rumba i jive en divuit esportistes de ball esportiu d’elit (edat mitjana: 19.6 ± 3.2 anys). Els mesuraments es van fer mitjançant electromiografia de superfície (EMG) durant l’execució de la coreografia. Es va registrar i es va analitzar l’EMG en ambdues cames del recte femoral (RF), el bíceps femoral (BF), el tibial anterior (TA) i el gastrocnemi medial (GM). En la rumba, l’activació total del RF (mitjana, 115.95; IQR, 36.00 mV) va ser menor que la del BF (mitjana, 146.68; IQR, 10.02 mV; p = .002) i el GM (mitjana, 149.81; IQR, 85.66 mV; p = .035). En el jive, l’activació global més gran va correspondre al BF (mitjana, 155.40; IQR, 44.89 mV), i les diferències van tenir significació estadística en comparació amb l’activació del TA (mitjana, 123.09; IQR, 51.24 mV; p = .028). Es van trobar diferències significatives entre la rumba i el jive en les fibres tipus I del RF (p ≤ .05), les fibres tipus IIa del TA (p ≤ .05); i les fibres tipus IIb del GM (p ≤ .05), tant en homes com en dones. Entre els ballarins, hi va haver diferències en les fibres de tipus IIb del GM (p ≤ .05), mentre que es van observar diferències en les fibres tipus I del TA entre les dones (p ≤ .05). Aquest estudi mostra evidències experimentals d’una activació muscular significativament diferent per a l’extremitat inferior en balls amb diferent tempo. Els resultats d’aquest estudi aporten informació rellevant per optimitzar els programes d’entrenament d’alt rendiment i prevenció de lesions, que és clau per a l’èxit professional en el ball esportiu.
Introducció
El ball esportiu és una combinació d’art, esport i rendiment esportiu. Es practica per parelles, la qual cosa permet expressar emocions i formar moviments harmoniosos en resposta a diferents tipus de música (Lukić et al., 2011; Riding et al., 2013; Uzunović et al., 2009; Uzunović i Kostić, 2005). Aquest esport es compon de tres especialitats: balls llatins, estàndards i els “10 balls”; aquests últims són una combinació de balls estàndards i llatins (WDSF, 2017). Els balls llatins es caracteritzen principalment per figures obertes, o semiobertes, i tancades que requereixen una connexió visual (Čačković et al., 2012). Els balls que conformen l’especialitat llatina són: la samba, amb un tempo de 50–52 pulsacions per minut (ppm); el txa-txa-txa, amb un tempo de 30–32 ppm; la rumba, amb un tempo de 25–27 ppm; el pasdoble, amb un tempo de 60–62 ppm, i el jive, amb un tempo de 42–44 ppm (FEBD, 2017).
La rumba és un ball amb ritme corporal molt expressiu en el qual es representen moviments atractius i delicats. La fluïdesa i el ritme del moviment als músculs de l’esquena i l’acció del maluc dels ballarins són de gran importància (Shang, 2013). D’aquesta manera, els malucs dibuixen de manera natural una trajectòria de moviment en un “8” invertit controlat pels ballarins (Shang, 2013).
El jive és el ball llatí més explosiu i provoca una alta freqüència cardíaca (Bria et al., 2011; Liiv et al., 2014). Fonamentalment, el jive es caracteritza per l’equilibri entre el SWINGy (amb gir) i el JUMPy (amb salt), dos principis bàsics que contribueixen al rendiment general del ball (Dance Comp Review, 2014).
El SWINGy és present en cada un dels passos executats en el jive. El cos s’inclina cap endavant, provocant un desplaçament lateral mitjançant el moviment del maluc, i es manté dret durant els passos cap endavant o cap enrere. El suport del peu s’aconsegueix principalment mitjançant el suport del primer i segon metatarsians (Dance Comp Review, 2014).
Quant al JUMPy, l’acció principal és la flexió del genoll simultani a la contracció de l’abdomen, que dona lloc a un petit salt. Es compon de quatre fases: pas, salt, vol i aterratge. En la primera fase, el pas es fa amb una petita flexió al genoll, la qual cosa dona lloc a la fase següent: l’extensió del genoll, que produeix l’impuls per saltar. Això es trasllada a la fase de vol i posteriorment a la d’aterratge, on la major part del suport el proporcionen els primers metatarsians amb una lleugera flexió dels genolls per absorbir l’impacte (Dance Comp Review, 2014).
L’electromiografia de superfície és una tècnica habitual i no invasiva per analitzar les contraccions musculars en aplicacions reals (Hermens i Freriks, 1997; Liu et al., 2002). En els éssers humans, les fibres ràpides i lentes no estan separades físicament, però l’evolució ha mantingut d’alguna manera la separació dels diferents tipus de fibres. Les unitats motrius del múscul humà se solen classificar en tres grups diferents comunament denominats: oxidatiu lent o tipus I (TI); oxidatiu ràpid o tipus IIa (TIIa); i glucolític ràpid o tipus IIb (TIIb) (Von Tscharner i Goepfert, 2006; Brooke i Kaiser, 1970). Aquests grups poden ser reclutats en diferents proporcions per a diferents períodes d’un moviment i poden explicar almenys part de la variabilitat espectral (Von Tscharner i Goepfert, 2003; Wakeling et al., 2001). Per analitzar les dades obtingudes amb l’electromiografia de superfície en contraccions musculars variables, s’han adoptat tècniques de mesurament de la freqüència temporal (Kumar et al., 2003). La transformada de Wavelet contínua (comparació de diferents tècniques de freqüència al llarg del temps) produeix resultats precisos amb una bona representació de la localització temporal i freqüencial (Karlsson et al., 2000).
Es disposa d’escasses proves sobre el ball esportiu i, en particular, sobre l’activitat muscular dels ballarins. Zagorc et al. (2010) van utilitzar la tensiomiografia per estudiar el temps de contracció d’esportistes de ball esportiu i van observar que el temps de contracció en músculs com el gastrocnemi variava entre gèneres. Liébana et al. (2017) van analitzar els EMG d’esportistes de ball esportiu que interpretaven la rumba bolero i van observar les diferències. D’aquesta manera, es van trobar diferències d’activació al recte femoral (RF) i el gastrocnemi medial (GM) de les dones associades amb el bolero-rumba, així com en el tibial anterior (TA) i el GM, a més de diferències entre els ritmes de diversos gèneres de ball (Haeufle et al., 2010).
Per comprendre com el complex sistema musculoesquelètic pot generar una força adequada a les cames, és necessari conèixer les propietats intrínseques dels músculs (Haeufle et al., 2010). Per això, la finalitat del nostre estudi era avaluar l’activitat muscular dels tres tipus de fibres musculars del RF, el BF, el TA i el GM dels ballarins, comparant la rumba amb el jive en homes i dones a fi de dissenyar entrenaments específics per a aquests esportistes. Els resultats previstos indicarien diferències significatives en l’activació dels tres tipus de fibres musculars analitzades: RF, BF, TA i GM (en ambdues cames, dominant i no dominant) en les extremitats inferiors dels ballarins, comparant la rumba amb el jive, i entre gèneres.
Materials i metodologia
Participants
Els participants van ser 18 esportistes de ball esportiu (nou parelles). Tots ells són ballarins de categoria A (la màxima categoria) amb 10.44 ± 3.51 anys d’experiència en el ball; a més, són especialistes en els “10 balls” o en balls llatins (vegeu la Taula 2).
Els criteris d’inclusió exigien que els participants estiguessin en actiu durant l’estudi, tinguessin més de divuit anys i s’ajustessin als 10 balls o a la modalitat llatina; els participants no havien d’haver patit lesions durant l’any anterior i havien d’haver ballat en la categoria A durant almenys un any amb la mateixa parella. Es va contactar amb sis escoles de dansa que podien estar interessades a participar en aquest estudi. D’un total de 10 parelles que complien els criteris d’inclusió, se’n van mesurar nou, és a dir, 18 participants, tots ells amb dominància dreta. Una parella va ser exclosa dels mesuraments per problemes d’agenda; a més, aquests ballarins van dissoldre posteriorment la parella.
Procediments
Es va demanar als ballarins que no fessin cap exercici físic en les 24 hores prèvies a la sessió de l’estudi. Durant la sessió, es van prendre mesures de l’estatura (amb un tallímetre SECA 709 7021994; Seca GmbH i Co. KG., Alemanya) i dades antropomètriques (pes i índex de massa corporal) mitjançant impedància bioelèctrica (analitzador de composició corporal segmentària Tanita BC-418 MA; Tanita Corporation, Japó). Per recopilar les dades amb electromiografia (EMG), es va seguir el protocol estàndard (vegeu la Taula 1) per preparar la pell dels participants i col·locar els elèctrodes (Torrence i Compo, 1998; Welch, 1967).

Posteriorment, els participants van fer una adaptació idèntica a l’escalfament RAMP, que va consistir a integrar la mobilitat articular en turmells, genolls, malucs i espatlles, seguida de moviments específics per parelles sense música, i van continuar l’activació mitjançant esquats i planxes; finalment, les parelles van fer un ball al ritme de la música (Jeffreys, 2007). Es van efectuar mesuraments mitjançant electromiografia de superfície de l’activació muscular durant totes les coreografies competitives (120 s cada tipus de ball), preparades i interpretades pels ballarins (Mega Electronics Ltd., Kuopio, Finlàndia).
Hi ha treballs previs amb coreografies preestablertes (Liébana et al., 2018), per la qual cosa aquest estudi pretén mesurar l’activació en un context ecològic, apropant-nos al context real de competició.
L’estudi va ser aprovat pel Comitè d’Ètica de la Recerca de la Universitat Catòlica de València Sant Vicent Màrtir, amb el codi UCV/2015-2016/60, i s’ajusta a la Declaració de Hèlsinki. Els participants coneixien la finalitat de l’estudi i tots van rebre un consentiment informat per escrit.
Tractament de dades
Tots els mesuraments d’EMG es van recollir amb sensors Mega WBA amb una freqüència de mostratge de 1,000 Hz, un pas de banda de freqüència del sensor entre 20 i 500 Hz, 200 elèctrodes d’escuma Kendall amb hidrogel adhesiu conductor (col·locats amb una distància màxima entre elèctrodes de 20 mm) i es van compilar utilitzant el programari Megawin 3.1 (Mega Electronics Ltd., Kuopio, Finlàndia). A continuació, es van transferir a un arxiu ASCII per a la seva anàlisi posterior. Les dades extretes de l’activació muscular s’expressen en mil·livolts (mV). L’arxiu es va transformar a .m per analitzar-lo mitjançant Matlab.
El tractament de les dades es va iniciar utilitzant Matlab R2017b, que selecciona automàticament els segons centrals de cada exercici. El senyal es va filtrar mitjançant l’ús d’un filtre passabanda per establir els valors mínims amb un límit de 20 Hz i valors màxims de 400 Hz. Es va obtenir l’arrel quadràtica mitjana (RMS). Es va fer una transformada de Fourier utilitzant la transformada ràpida de Fourier (Welch, 1967), que indica l’espectre de potència mitjana i permet estimar la densitat espectral. Per a això, es va fer servir el periodograma de Welch amb una finestra de Hamming de longitud 1024, a fi d’estimar la densitat espectral (Welch, 1967). En aquest mètode, es duu a terme la fragmentació de la sèrie temporal, calculant d’aquesta manera un periodograma modificat per a cada un dels segments. Una vegada calculada la mitjana, aquest procés facilita l’estimació de la densitat espectral. El mètode Welch és una millora del mètode estàndard del periodograma, ja que efectua una reducció del soroll en l’espectre de potència estimat. Tot i així, aquest mètode planteja un problema. Per corregir-lo, es va aplicar una anàlisi de temps-freqüència, en el qual una finestra de longitud fixa es desplaça al llarg del senyal per relacionar les freqüències amb el temps i es poden avaluar aquestes freqüències a cada finestra.
A continuació, s’aplica una anàlisi de sèries temporals no estacionàries mitjançant la transformada de Wavelet (Torrence i Compo, 1998), un mètode que permet analitzar el senyal en el domini de l’escala temporal. Es tracta d’una sèrie temporal formada per famílies de funcions definides temporalment i espacialment, que es produeixen per escalat i translació d’una funció denominada “funció base”. L’escalograma consisteix en un espectre de potència amitjanat per a les diferents freqüències o escales, concedides en cada valor temporal (Torrence i Compo, 1998).
La transformada de Wavelet es divideix en dues variables: la transformada de Wavelet contínua detecta patrons o modificacions al llarg de l’evolució temporal del senyal a diferents escales, mentre que la transformada de Wavelet discreta s’obté mitjançant la descomposició del senyal en diferents zones de l’espectre de freqüències, seguit del filtratge de dades per obtenir els coeficients de Wavelet. El filtratge es produeix en relació amb l’aproximació, el detall i els filtres de pas baix (5 Hz) i pas alt (250 Hz). Aquests resultats són la descomposició del senyal global en senyals ortogonals que permeten dividir els senyals en cada una de les bandes de freqüència. En aquest cas, es van fixar tres bandes: la primera, < 70 Hz; la segona, entre 70 i 125 Hz, i l’última, entre 126 i 250 Hz (Torrence i Compo, 1998).
Anàlisi estadística
Per analitzar les dades, es va utilitzar el paquet estadístic SPSS 22.0 (IBM, Chicago, Illinois, EUA). Les característiques descriptives de l’antropometria es presenten com a mitjanes i desviacions típiques (DT). A causa de la mida limitada de la mostra (9 parelles), es van recomanar proves no paramètriques per comparar les variables quantitatives. Els canvis en l’activació muscular entre les dues modalitats de dansa es van avaluar amb la prova de rangs de Wilcoxon. Atesa la possible variabilitat dels mesuraments d’EMG en els participants, els valors es van presentar com a mitjanes i rangs interquartílics (IQR). Les comparacions de variables quantitatives entre ballarins i ballarines es van avaluar amb la prova de Mann-Whitney. També es va indicar el valor Z. Per a tots els efectes principals i interaccions, es va adoptar un nivell de confiança de .05.
Resultats
Les característiques antropomètriques descriptives de la mostra es presenten a la Taula 2. No hi va haver diferències significatives entre ballarins i ballarines en les variables antropomètriques (Taula 2).
Considerant l’activació en l’EMG de totes les fibres musculars juntes en els diferents músculs analitzats, la rumba va mostrar menys activació que el jive als músculs de la part proximal de l’extremitat inferior (RF i BF), encara que no hi va haver diferències entre les dues modalitats de ball (Figura 1A). En la rumba, l’activació del RF (mitjana, 115.95; IQR, 36.00 mV) va ser inferior que la del BF (mitjana, 146.68; IQR, 10.02 mV; p = .002) i el GM (mitjana, 149.81; IQR, 85.66 mV; p = .035). El GM va mostrar l’activació muscular més gran en la rumba. En el jive, l’activació més gran va correspondre al BF (mitjana, 155.40; IQR, 44.89mV), i les diferències van tenir significació estadística en comparació amb l’activació del TA (mitjana, 123.09; IQR, 51.24mV; p = .028). No hi va haver diferències entre la cama dominant i la no dominant en l’activació en l’EMG global dels músculs durant les dues modalitats de dansa.
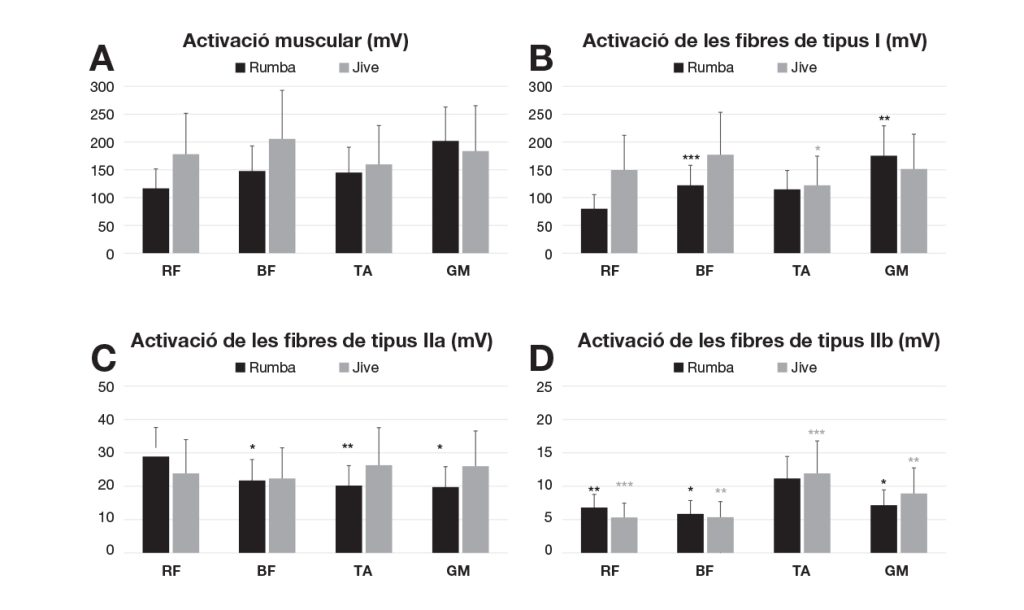
Nota: *p < .05; **p < .01; ***p = .001. A: registre mitjançant EMG de l’activació global dels diferents músculs analitzats a la cama dominant; B: activació de les fibres de tipus I; C: activació de les fibres de tipus IIa; i D: activació de les fibres de tipus IIb a la cama dominant. (RF: Recte femoral; BF: Bíceps femoral; TA: Tibial anterior; GM: Gastrocnemi medial).
A la figura 1, B, C i D mostren una comparació més detallada entre l’activació de diferents tipus de fibres en els músculs avaluats a la cama dominant en relació amb la modalitat de dansa. En la rumba, l’activació de les fibres de tipus I del RF va ser significativament menor que en el BF (z = -3.201; p = .001) i GM (z = -2.635; p = .008). A més, el GM va mostrar l’activació muscular més gran de fibres de tipus I en la rumba. En el jive, l’activació més gran va correspondre al BF (mitjana, 124.05; IQR, 55.35 mV), i les diferències van tenir significació estadística en comparació amb l’activació del TA (mitjana, 85.90; IQR, 35.52 mV; p = .028).
Quant a les fibres de tipus IIa, el múscul RF va mostrar una activació més gran en la rumba que en el jive. En la rumba, l’activació del RF va ser significativament més gran que la del BF (z = -2.3301; p = .020), el TA (z = -2.809; p = .005) i el GM (z = -2.243; p = .025) (Fig. 1C).
L’activació de les fibres de tipus IIb va ser molt baixa a tots els músculs estudiats. En ambdues modalitats de dansa, l’activació més gran es va trobar en el TA (Figura 1D). En la rumba, l’activació del TA va mostrar diferències amb significació estadística en comparació amb el RF (z = -2.940; p. = 0,003), el BF (z = -2.461; p = .014), i el GM (z = -2.025; p = .043). En el jive, l’activació del TA també va mostrar diferències amb significació estadística en comparació amb el RF (z = -3.201; p = .001), i el BF (z = -3.157; p = .002). En el jive, també hi va haver diferències entre l’activació del RF i del GM (z = -2.765; p = .006), i entre el BF i el GM (z = -3.157 p = .002).
A la Taula 3, es presenten les dades descriptives sobre l’activació dels diferents tipus de fibres musculars, tant a la cama dominant com en la no dominant, durant les dues modalitats de dansa. Quant a les fibres musculars de tipus I, la rumba va mostrar menys activació que el jive als músculs RF i BF de la cama dominant, però les diferències només van ser significatives per al RF (p = .022). En les fibres de tipus IIa, el TA i el GM de la cama dominant van mostrar més activació en el jive que en la rumba, però les diferències només van tenir significació estadística en el TA (p = .002). L’activació de les fibres de tipus IIb va ser molt baixa en tots els músculs estudiats. Les diferències entre la rumba i el jive només es van detectar en el GM de la cama no dominant (p = .016). Quan es van comparar les cames dominants i no dominants dins de cada modalitat de ball, només hi va haver diferències en l’activació de les fibres de tipus I del GM, que van ser més grans a la cama dominant en la rumba (p = .006). En el jive, no hi va haver diferències en l’activació dels diferents músculs analitzats. Els músculs distals (TA i GM) es van activar més en el jive que en la rumba. En el jive, l’activació més gran de fibres de tipus IIa es va detectar en el GM en tots dos costats (Taula 3).
A la Taula 4, es mostren les dades descriptives relatives als tipus de fibres per a la rumba i el jive en tots dos sexes. Es van obtenir diferències significatives per a homes i dones en les fibres tipus I del TA, tant en la rumba com en el jive, a la cama dominant (p = .028). En les dones, hi va haver diferències significatives en l’activació no dominant del TA en la rumba i el jive (p = .038). Quan es van comparar ballarins i ballarines dins de cada modalitat, només es van trobar diferències en l’activació EMG en el GM de la cama dominant durant la rumba (Taula 4). Els homes van mostrar una activació inferior de les fibres de tipus I en el GM (p = .019) i una activació més gran de les fibres de tipus IIa en el GM (p = .014) que les dones.
Discussió
Els ballarins en general, i particularment el ball esportiu, no s’han estudiat en profunditat i existeix una notable falta d’informació sobre el comportament de les fibres musculars durant la pràctica d’aquest esport. La importància del present estudi rau en les diferències significatives identificades entre la rumba i el jive, les quals faciliten la programació i planificació de l’entrenament neuromuscular segons els nostres resultats, si bé som conscients de les limitacions degudes al nombre reduït de la mostra, ja que es tracta d’un esport minoritari. Aquests balls presenten diferències importants quant a tècnica, ritme i tempo, però el fet que es produeixin clares diferències en l’activació muscular també s’ha de tenir en compte en el desenvolupament de les sessions d’entrenament per optimitzar el rendiment i reduir el risc de lesions.
La tècnica, el ritme i els moviments de la rumba són més lents que en el jive. En la rumba, es busca una estètica particular, per la qual cosa la seva tècnica afavoreix l’extensió de les extremitats inferiors (Shang, 2013). En canvi, el jive té característiques similars als salts a peu coix, amb les seves puntades de peu i salts consecutius. Aquest exercici combina velocitat i força per produir un moviment explosivo-reactiu (Cappa i Behm, 2013). Aquests exercicis impliquen contraccions musculars excèntriques (estirament) i concèntriques (escurçament), generalment utilitzant el cos com a sobrecàrrega i generant un cicle d’estirament-escurçament (SSC) (Cappa i Behm, 2013). Perquè una acció muscular es classifiqui com a cicle d’estirament-escurçament durant l’activitat de rebot, el patró d’activació muscular ha d’incloure una preactivació prèvia al contacte amb el terra, una acció excèntrica ràpida i una transició immediata i ràpida entre les fases excèntrica i concèntrica (Komi, 2000). Per aquesta raó, s’observen diferències significatives entre la rumba i el jive en RF1_TI, TA1_TIIa i GM2_TIIb a la cama no dominant. De la mateixa manera que en els salts a peu coix, en el jive els músculs isquiotibials i els quàdriceps han de ser actius alhora per crear estabilitat (Wibawa et al., 2016).
El TA és un múscul que s’activa no només en la dorsiflexió del peu, sinó també per controlar la pronació, per la qual cosa en el jive estaria actuant excèntricament per contribuir a tots dos moviments (Cappa i Behm, 2013). Per tant, les diferències entre la rumba i el jive es troben principalment en el TA1_TIIa. Les fibres del TA1_TIIa s’activen en moviments curts i d’alta intensitat, ja que aquest múscul proporciona suport mitjançant l’activació excèntrica de frenada. Aquesta activació també es veu afavorida per al suport i quan els ballarins executen una tècnica d’aterratge utilitzant el primer i segon metatarsians (Dance Comp Review, 2014).
També es van trobar diferències significatives entre la rumba i el jive en relació amb el GM2_TIIb. Això pot ser degut al treball d’SSC d’aquest múscul, ja que una tècnica de jive afavoreix la realització contínua de salts ràpids i puntades de peu, que genera treball pliomètric i la realització d’un cicle d’estirament-escurçament (Cappa i Behm, 2013). Nicol et al. (2006) destaquen en el seu treball que el gastrocnemi reacciona de manera diferent en l’aterratge després d’un salt. Si el salt és petit, les fibres del gastrocnemi tendeixen a escurçar-se per frenar. Si el salt és alt i requereix una frenada important, les fibres musculars tendeixen a allargar-se. Això es deu al fet que es produeix menys resistència a l’estirament pel possible alliberament de ponts transversals (Nicol et al., 2006). La càrrega d’impacte determina el comportament del fascicle en un múscul específic i la intensitat de l’esforç després de la fase de frenada té certa influència en aquesta interacció atès que afecta el retrocés del tendó en l’impuls final (Nicol et al., 2006).
D’altra banda, els nostres resultats mostren diferències en l’activació muscular entre homes i dones. Aquestes dades estarien en consonància amb les obtingudes en l’estudi de Liébana et al. (2017). Es van observar discrepàncies en l’activació muscular entre sexes en el tibial anterior i el gastrocnemi, la qual cosa demostra que l’activació muscular entre homes i dones és diferent (Haeufle et al., 2010). Això es pot deure tant a les diferències entre els passos de les dones i els dels homes com a la marcada diferència entre els talons de les sabates de ball d’homes i dones que, en conseqüència, provoca que els músculs generin forces actives diferents a les cames. Hill (1938) va descriure i va distingir les propietats intrínseques d’un sol múscul, representat per un element elàstic en sèrie i un element contràctil amb relacions força-longitud i força-velocitat. Les propietats musculars poden compensar les pertorbacions i facilitar la convergència dels moviments dinàmics i explosius. Les propietats intrínseques del múscul representades per la funció força-longitud-velocitat en els models musculars de Hill actuen com un sistema de retroalimentació perifèrica sense retard (Haeufle et al., 2010).
Les diferències en l’activació muscular entre els tipus de fibres, músculs, sexes i balls observats en el nostre estudi demostrarien la necessitat d’un entrenament individualitzat i planificat per a cada tipus de ball i parella (home davant dona). D’allà la necessitat d’un entrenament específic per a ballarins que abordi el tipus de força treballada, l’entrenament de la mobilitat, la tècnica i el control motor. Aquests factors són de gran importància per prevenir lesions i aconseguir el màxim rendiment esportiu.
En relació amb les dades mostrades en aquest estudi, es poden destacar les limitacions quant a la mostra, ja que es tracta d’un nombre baix de participants en comparació amb altres estudis, però representatiu del nombre de participants en el campionat d’Espanya, tenint en compte que el ball esportiu és un esport minoritari.
Conclusions
Els resultats d’aquest estudi proporcionen dades fàctiques experimentals d’activacions musculars significativament diferents per a l’extremitat inferior en funció del sexe i la modalitat de dansa. Es van trobar diferències significatives en l’activació en funció del tipus de fibra entre la rumba i el jive. Des d’una perspectiva clínica, les nostres troballes poden ajudar els entrenadors i metges esportius a comprendre el perfil esportiu específic dels ballarins d’elit. El coneixement de la funció muscular, l’activitat i l’equilibri és crucial per optimitzar l’alt nivell de rendiment d’aquests esportistes, així com per afavorir els programes de prevenció de lesions, que són crucials per maximitzar el seu èxit esportiu. Aquests resultats suposen un primer pas per proporcionar valors de referència de les fibres musculars implicades en els moviments dels esports de dansa; aquests valors poden contribuir a dissenyar exercicis que afavoreixin el rendiment esportiu i la prevenció de lesions. En fer aquest tipus d’anàlisi mitjançant transformades de Wavelet, s’ha pogut observar que aquesta anàlisi és sensible i mostra resultats coherents, tenint en compte a tota hora les limitacions de l’estudi. Es poden utilitzar transformades de Wavelet per analitzar moviments amb característiques similars (Cappa i Behm, 2013). Aquesta anàlisi permet comprendre les exigències musculars dels diferents tipus de dansa. Per tant, per al desenvolupament correcte del jive és important el treball pliomètric, amb un cicle d’escurçament-estirament. Així mateix, per minimitzar el desequilibri muscular entre la cadena posterior i l’anterior, així com la asimetria entre la cama dominant i la no dominant, es recomanaria el treball de força.
Referències
[1] Bria, S., Bianco, M., Galvani, C., Palmieri, V., Zeppilli, P., & Faina, M. (2011). Physiological characteristics of elite sport-dancers. The Journal of Sports Medicine and Physical Fitness, 51(2), 194-203.
[2] Brooke, M. H., & Kaiser, K. K. (1970). Muscle Fiber Types: How Many and What Kind? Archives of Neurology, 23(4), 369-379. doi.org/10.1001/archneur.1970.00480280083010
[3] Čačković, L., Baric, R., & Vlasic, J. (2012). Psychological stress in dancesport. Acta Kinesiologica, 6(2), 71-74. actakin.com/PDFS/BR0602/SVEE/04%20CL%2013%20LC.pdf
[4] Cappa, D. F., & Behm, D. G. (2013). Neuromuscular Characteristics of Drop and Hurdle Jumps With Different Types of Landings: Journal of Strength and Conditioning Research, 27(11), 3011-3020. doi.org/10.1519/JSC.0b013e31828c28b3
[5] Criswell, E., & Cram, J. R. (Eds.). (2011). Cram’s introduction to surface electromyography (2nd ed). Jones and Bartlett.
[6] Dance Comp Review. (2014). Principals of International Latin Jive. Dance Comp Review. dancecompreview.com/principals-international-latin-jive/
[7] FEBD. (2017). Código Básico de Competiciones. www.febd.es/wordpress/wp-content/uploads/2017/01/codigo_basico_de_competiciones_11_5.pdf
[8] Haeufle, D. F. B., Grimmer, S., & Seyfarth, A. (2010). The role of intrinsic muscle properties for stable hopping—Stability is achieved by the force–velocity relation. Bioinspiration & Biomimetics, 5(1), 016004. doi.org/10.1088/1748-3182/5/1/016004
[9] Hermens, H., & Freriks, B. (1997). The state of the art on sensors and sensor placement procedures for surface electromyography: A proposal for sensor placement procedures. Roessingh Research and Development.
[10] Hill, A. V. (1938). The Heat of Shortening and the Dynamic Constants of Muscle. Proceedings of the Royal Society B: Biological Sciences, 126(843), 136-195. doi.org/10.1098/rspb.1938.0050
[11] Jeffreys, I. (2007). Warm up revisited – the ‘ramp’ method of optimising performance preparation. Professional Strength and Conditioning, 6, 12-18.
[12] Karlsson, S., Jun Yu, & Akay, M. (2000). Time-frequency analysis of myoelectric signals during dynamic contractions: A comparative study. IEEE Transactions on Biomedical Engineering, 47(2), 228-238. doi.org/10.1109/10.821766
[13] Komi, P. V. (2000). Stretch-shortening cycle: A powerful model to study normal and fatigued muscle. Journal of Biomechanics, 33(10), 1197-1206. doi.org/10.1016/S0021-9290(00)00064-6
[14] Kumar, D. K., Pah, N. D., & Bradley, A. (2003). Wavelet analysis of surface electromyography. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 11(4), 400-406. doi.org/10.1109/TNSRE.2003.819901
[15] Liébana, E., Blasco, E., Monleón, C., Pablos, C., & Moratal, C. (2017). Muscular activation in rumba bolero in elite dancers of DanceSport. Journal of Human Sport & Exercise, 12(3proc), S807-S812. doi.org/10.14198/jhse.2017.12.Proc3.04
[16] Liébana, E., Monleón, C., Morales, R., Pablos, C., Moratal, C., & Blasco, E. (2018). Muscle Activation in the Main Muscle Groups of the Lower Limbs in High-Level Dancesport Athletes. Medical Problems of Performing Artists, 33(4), 231-237. doi.org/10.21091/mppa.2018.4034
[17] Liiv, Jürimäe, T., Mäestu, J., Purge, P., Hannus, A., & Jürimäe, J. (2014). Physiological characteristics of elite dancers of different dance styles. European Journal of Sport Science, 14, S429-S436. doi.org/10.1080/17461391.2012.711861
[18] Liu, J. Z., Brown, R. W., & Yue, G. H. (2002). A dynamical model of muscle activation, fatigue, and recovery. Biophysical Journal, 82(5), 2344-2359. www.ncbi.nlm.nih.gov/pmc/articles/PMC1302027/
[19] Lukić, A., Bijelić, S., Zagorc, M., & Zuhrić-Sebić, L. (2011). The importance of strength in sport dance performance technique. Sportlogia, 7(1), 61-67. doi.org/10.5550/sgia.110701.en.061L
[20] Nicol, C., Avela, J., & Komi, P. V. (2006). The Stretch-Shortening Cycle. Sports Medicine, 36(11), 977-999. doi.org/10.2165/00007256-200636110-00004
[21] Riding, T., Wyon, M., Ambegaonkar, J., & Redding, E. (2013). A Bibliographic Review of Medicine and Science Research in DanceSport. Medical Problems of Performing Artists, 28(2), 70-79. doi.org/10.21091/mppa.2013.2013
[22] Shang, Y. (2013). Technical analysis of the hips squeezing action in rumba based on biomechanics. Trade Science Inc, 8(9), 1205-1209.
[23] Torrence, C., & Compo, G. P. (1998). A practical guide to wavelet analysis. Bulletin of the American Meteorological Society, 79(1), 61-78. journals.ametsoc.org/doi/abs/10.1175/1520-0477(1998)079%3C0061:APGTWA%3E2.0.CO;2
[24] Uzunović, S., & Kostić, R. (2005). A study of success in A study of success in Latin American sport dancing. Facta Universitatis, Series Physical Education and Sport 3, 23-35.
[25] Uzunović, S., Kostić, R., & Miletić, Đ. (2009). Motor status of competitive young sport dancers-gender differences. Acta Kinesiologica, 3(1), 83-88. www.actakin.com/PDFS/BR0301/SVEE/04%20CL%2014%20SU.pdf
[26] Von Tscharner, V., & Goepfert, B. (2003). Gender dependent EMGs of runners resolved by time/frequency and principal pattern analysis. Journal of Electromyography and Kinesiology, 13(3), 253-272. doi.org/10.1016/S1050-6411(02)00111-6
[27] Von Tscharner, V., & Goepfert, B. (2006). Estimation of the interplay between groups of fast and slow muscle fibers of the tibialis anterior and gastrocnemius muscle while running. Journal of Electromyography and Kinesiology, 16(2), 188-197. doi.org/10.1016/j.jelekin.2005.07.004
[28] Wakeling, J. M., Pascual, S. A., Nigg, B. M., & Tscharner, V. (2001). Surface EMG shows distinct populations of muscle activity when measured during sustained sub-maximal exercise. European Journal of Applied Physiology, 86(1), 40-47. doi.org/10.1007/s004210100508
[29] WDSF. (2017). World Dance Sport Federation. About DanceSport. www.worlddancesport.org/about
[30] Welch, P. (1967). The use of fast Fourier Transform for the Estimation of Power Spectra: A Method based on time avering over short, modilied periodograms. 15, 70-73. doi.org/10.1109/TAU.1967.1161901
[31] Wibawa, A. D., Verdonschot, N., Halbertsma, J. P. K., Burgerhof, J. G. M., Diercks, R. L., & Verkerke, G. J. (2016). Musculoskeletal modeling of human lower limb during normal walking, one-legged forward hopping and side jumping: Comparison of measured EMG and predicted muscle activity patterns. Journal of Biomechanics, 49(15), 3660-3666. doi.org/10.1016/j.jbiomech.2016.09.041
[32] Zagorc, M., Šimunič, B., Pišot, R., & Oreb, G. (2010). A comparison of contractile parameters among twelve skeletal muscles of inter-dance couples. Kinesiologia Slovenica, 16(3), 57-65.
ISSN: 2014-0983
Rebut: 2 de juny de 2023
Acceptat: 10 d'octubre de 2023
Publicat: 1 d'abril de 2024
Editat per: © Generalitat de Catalunya Departament de la Presidència Institut Nacional d’Educació Física de Catalunya (INEFC)
© Copyright Generalitat de Catalunya (INEFC). Aquest article està disponible a la url https://www.revista-apunts.com/. Aquest treball està publicat sota una llicència Internacional de Creative Commons Reconeixement 4.0. Les imatges o qualsevol altre material de tercers d’aquest article estan incloses a la llicència Creative Commons de l’article, tret que s’indiqui el contrari a la línia de crèdit; si el material no s’inclou sota la llicència Creative Commons, els usuaris hauran d’obtenir el permís del titular de la llicència per reproduir el material. Per veure una còpia d’aquesta llicència, visiteu https://creativecommons.org/licenses/by/4.0/deed.ca


