El área de histéresis de las variables psicobiológicas. ¿Nuevo biomarcador no invasivo de la acumulación de esfuerzo?
Lluc Montull

Óscar Abenza
Robert Hristovski
Natàlia Balagué
*Correspondencia: Natàlia Balagué nbalague@gencat.cat
Citación
Montull, L., Abenza, O., Hristovski, R. & Balagué, N. (2023). Hysteresis Area of Psychobiological Variables. A New Non-Invasive Biomarker of Effort Accumulation? Apunts Educación Física y Deportes, 152, 55-61. https://doi.org/10.5672/apunts.2014-0983.es.(2023/2).152.06
Resumen
El área de histéresis, que refleja la dependencia de los parámetros psicobiológicos respecto a los antecedentes, se ha sugerido recientemente como nuevo marcador no invasivo del estrés y la tolerancia en relación con el ejercicio. Sin embargo, aún no se han explorado adecuadamente sus prometedoras aplicaciones. El objetivo de este estudio es comprobar si el área de histéresis del esfuerzo percibido (TEP), la frecuencia cardiaca (FC) y la concentración de oxihemoglobina muscular (O2Hb muscular) son sensibles a la acumulación de esfuerzo a lo largo de series múltiples de ejercicio de carrera. Diez estudiantes de Educación Física realizaron cinco series consecutivas de carrera a velocidad aeróbica máxima. Las series consecutivas comenzaban cuando los participantes comunicaban al menos una TEP ≤ 11 (escala de 6 a 20 de Borg). Se calcularon las áreas de histéresis de la TEP, la FC y la O2Hb muscular para cada serie y se comparó su magnitud utilizando la prueba ANOVA de Friedman y la prueba de Wilcoxon. Los tamaños del efecto se calcularon mediante la d de Cohen. Las áreas de histéresis de todas las variables estudiadas cambiaron entre las series iniciales y finales: la TEP y la FC aumentaron (entre las series 1-4): Z = –1.99, p = .04; Z = –2.19, p = .03, respectivamente), y la O2Hb muscular disminuyó (entre las series 1-4: Z = –2.80, p < .01). Se observaron efectos de gran tamaño (d = 1.22 – 1.81) en la FC entre las series 1-4, 1-5, 2-4, 2-5, y en la O2Hb muscular entre las series 1-4 y 1-5. Los resultados mostraron que las áreas de histéresis de la TEP, la FC y la O2Hb muscular eran sensibles a la acumulación de esfuerzo durante ejercicios repetidos de carrera realizados a velocidad aeróbica máxima. Si bien es necesario seguir investigando, el área de histéresis de las variables psicobiológicas apunta a ser un biomarcador sensible para la supervisión de la fatiga aguda.
Introducción
En biología, el fenómeno de la histéresis se refiere a la dependencia del sistema objeto de estudio respecto a los antecedentes (es decir, la dependencia de la trayectoria), sello distintivo de su compleja naturaleza. Se caracteriza por una dinámica autoorganizada y no lineal (Hristovski et al., 2010, 2014) y, a menudo, se pasa por alto en la supervisión del ejercicio y la ciencia del ejercicio (Balagué et al., 2020). Explica el modo en que el parámetro estudiado vuelve a su estado inicial con cierto retraso tras verse alterado y por qué pueden coexistir varios estados de este parámetro para un mismo valor cuantitativo. Por ejemplo, puede observarse el mismo valor de concentración de ácido láctico en dos estados fisiológicos diferentes: durante el ejercicio y en reposo (pongamos durante la recuperación tras el ejercicio). El fenómeno se produce porque el sistema depende de sus antecedentes; en consecuencia, las trayectorias de comportamiento mayoritario de la variable estudiada son diferentes cuando el parámetro de control está modificándose sistemáticamente en direcciones opuestas (Hristovski et al., 2014).
El fenómeno de la histéresis de diversos parámetros se ha estudiado en diversos ámbitos relacionados con la salud, como la fisiología (Liu et al., 2014), la biomecánica (Butler et al., 1978) y la medicina (Cabasson et al., 2012). Recientemente, el comportamiento de histéresis de las variables psicobiológicas se ha propuesto como marcador no invasivo del estrés y la tolerancia en relación con la carga de trabajo del ejercicio (Montull et al., 2020) y se ha aplicado para estudiar los efectos del desentrenamiento de los programas de entrenamiento estandarizados (Martín-Guillaumes et al., 2021). Sin embargo, sus aplicaciones y posibilidades de supervisión aún no se han explorado adecuadamente.
La histéresis de una variable específica puede cuantificarse a través del área de histéresis, que refleja la cantidad de energía disipada tras la recuperación de la variable (Mayergoyz, 2003). La Figura 1 representa el área de histéresis correspondiente al espacio entre las fases creciente y decreciente de un protocolo de esfuerzo piramidal. Montull et al. (2020) demostraron que un área de histéresis más baja de la tasa de esfuerzo percibido (TEP) y la frecuencia cardiaca (FC) se asocia con una mayor pericia del deportista y una mayor eficacia de recuperación. En consecuencia, los autores sugirieron que el área de histéresis de las variables psicobiológicas también puede ser sensible a la acumulación de esfuerzo, que apenas se capta a través de las técnicas de análisis de datos y los protocolos de evaluación que se aplican habitualmente hoy en día (Halson, 2014).
Considerando la fatiga un producto de interacciones en red multidimensionales, multiescalares y dependientes del entorno, los valores cuantitativos fijos de variables aisladas o los diversos indicadores atemporales que suelen proporcionarse para definirla podrían resultar insuficientes, y a menudo imprecisos, para registrarla (Balagué et al., 2020). Esta evaluación atemporal, basada en una concepción excesivamente simplificada y distorsionada del organismo como un sistema adaptativo complejo (SAC), no tiene en cuenta la dinámica no lineal y fluctuante del comportamiento de los deportistas ni la idiosincrasia de sus respuestas dinámicas al ejercicio (Nesselroade y Molenaar, 2010). Por este motivo, el marco de la fisiología en red del ejercicio promueve la evaluación de propiedades de los SAC tales como la histéresis y el uso del análisis de series temporales individuales para evaluar los estados fisiológicos (Balagué et al., 2020).
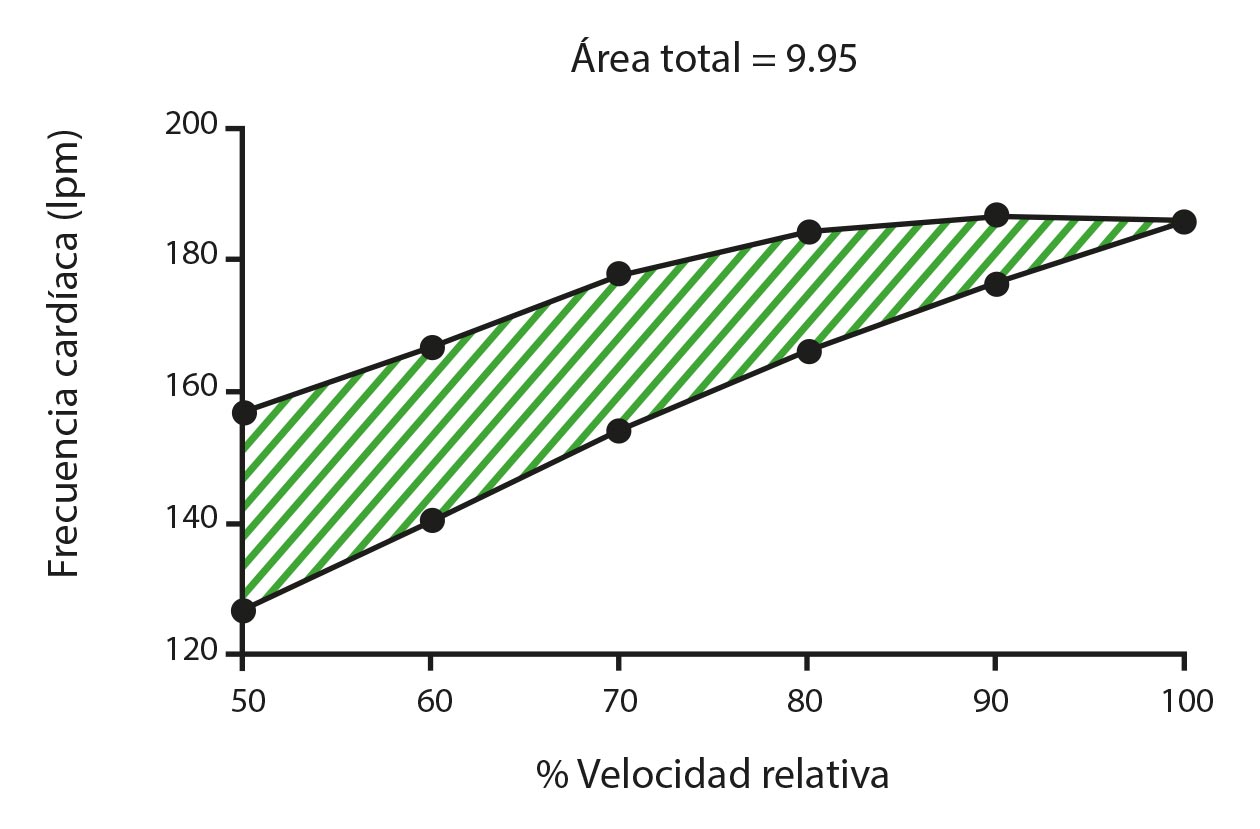
El desconocimiento de la respuesta de histéresis en determinados tipos de entrenamiento, como el entrenamiento por intervalos, puede llevar a una prescripción inadecuada de series de ejercicio y tiempos de recuperación. Las series de ejercicio suelen basarse en intensidades preestablecidas y valores psicobiológicos fijos (por ejemplo, la FC correspondiente al umbral anaeróbico) y los tiempos de recuperación, en hipótesis fijas basadas en el metabolismo (por ejemplo, el sistema de fosfágeno tarda de 2 a 5 min en recargarse) (Wilmore et al., 2008).
Queda mucho por conocer sobre la respuesta de histéresis al ejercicio. Tan solo se han estudiado la TEP, la FC y algunas variables relacionadas que se han analizado para la detección de enfermedades cardiovasculares (Cabasson et al., 2012). El área de histéresis de la saturación de oxígeno muscular (O2Hb muscular) (Hamaoka et al., 2011) podría arrojar luz sobre el modo en que los SAC compensan a escala microscópica el aumento de la demanda que conlleva la acumulación de esfuerzo.
En vista de lo aquí expuesto, el objetivo de este estudio era comprobar la sensibilidad del área de histéresis de la TEP, la FC y la O2Hb muscular a la acumulación de esfuerzo durante series múltiples de ejercicio de carrera. La hipótesis es que las áreas de la TEP y la FC aumentarán a lo largo de las series, mientras que las áreas de la O2Hb muscular disminuirán, ya que el esfuerzo acumulado reduce la saturación de oxígeno.
Metodología
Participantes
El estudio contó con la participación voluntaria de diez estudiantes de Ciencias del Deporte (cinco hombres y cinco mujeres: 20.50 ± 2.22 años; 1.74 ± 0.08 m; 64.25 ± 11.57 kg), que hacían ejercicio entre cinco y ocho horas/semana y llevaban una vida saludable y activa. Para determinar el tamaño de la muestra, se utilizó un tamaño del efecto grande ρ = 1, α = .05 y potencia (1-β) = .85. Se explicaron todos los procedimientos experimentales a los participantes antes de que dieran su consentimiento por escrito para participar. El experimento fue aprobado por el Comité de ética de investigaciones clínicas de la Administración deportiva de Cataluña (072015CEICEGC) y se llevó a cabo de acuerdo con la Declaración de Helsinki.
Protocolos de ensayo
Prueba de velocidad aeróbica máxima (VAM)
Se realizó una prueba de carga creciente y máxima en cinta rodante (h/p Cosmos Pulsar 3p®) para obtener la VAM de cada participante. La prueba comenzó a 8 km/h para los hombres y a 6 km/h para las mujeres, y la velocidad se incrementó en 1 km/h cada 100 s hasta que no pudieron mantener la velocidad impuesta. La velocidad final mantenida a lo largo de los 100 s se consideró la VAM. Esta prueba también se utilizó para familiarizar a los participantes con la escala de TEP de 6 a 20 de Borg.
Prueba de series múltiples
Una semana después de hacer la prueba de VAM, realizaron la prueba de series múltiples. Tras un calentamiento de 5 minutos a 8 km/h, los participantes realizaron cinco series consecutivas de 100 s al 100 % de la VAM (se recomiendan entre 30 y 120 s para las pruebas de capacidad anaeróbica láctica, Medbo y Tabata, 1993). Descansaron entre series hasta que registraron una TEP ≤ 11 (leve) en una escala de 6 a 20 de Borg. Tras cada serie, respondieron a la siguiente pregunta: «¿Cuál es tu esfuerzo percibido en este momento?». Se colocó la escala de TEP de 6 a 20 de Borg en un lugar visible.
Para evitar manipulaciones de los periodos de recuperación, no se informó a los participantes del número de series que debían realizar. Durante la prueba, no podían hablar. Al final, se les preguntó por sus sensaciones y podían hacer preguntas.
Obtención de datos
Durante el ejercicio, se supervisaron continuamente la FC (Polar Electro Oy, Finlandia) y la O2Hb muscular (PortaMon, Artinis, Medical System), por ser una de las variables más directas de la saturación de oxígeno en los músculos (McManus et al., 2018). En el vasto externo del cuádriceps de los participantes, se colocó el PortaMon (dispositivo de espectroscopia de frecuencias cercanas al infrarrojo), que registra a una frecuencia de muestreo de 10 Hz (McManus et al., 2018). Los valores se recogieron cada 25 s, al mismo tiempo que se registraba la TEP.

Análisis de los datos
Cálculo de las áreas de histéresis
Las áreas de histéresis de las variables psicobiológicas estudiadas se calcularon según Montull et al. (2020). Se tuvo en cuenta el espacio entre las fases de ejercicio y recuperación para cada variable y serie. Dado que la carga de trabajo era constante en todas las series de ejercicio y nula durante la recuperación, el parámetro de control seleccionado fue la duración del esfuerzo-recuperación (τ). Correspondía al tiempo actual de ejercicio o recuperación dividido entre el tiempo total de ejercicio (100 s). Las variables estudiadas y la τ se normalizaron de 0 a 10 para relativizarlas en la misma escala antes de calcular las áreas. Calculamos el área de histéresis total como la suma de las áreas parciales positivas y negativas. Las áreas se consideraron positivas cuando los valores de la fase de recuperación eran superiores a los de la fase de ejercicio, mientras que se consideraron negativas cuando los valores de la fase de recuperación eran inferiores a los de la fase de ejercicio (véase la Figura 2 a modo de ejemplo). En caso de que la fase de recuperación fuera más larga que la de ejercicio, aplicamos la misma lógica: hasta que los valores de recuperación no fueran inferiores al valor inicial de ejercicio, el área no se consideró negativa. Para este cálculo, se utilizó Matlab R2020a.
Análisis estadístico
Tras demostrar una distribución no normal, se aplicó la prueba ANOVA de Friedman y, a continuación, la prueba de Wilcoxon con el fin de comparar las diferencias de a) las áreas de histéresis entre todas las series (para cada variable), b) los valores iniciales de las variables entre series consecutivas, y c) las áreas de histéresis entre las tres variables estudiadas. También se calculó la d de Cohen para las áreas de histéresis y los valores iniciales de cada variable entre las series iniciales (1-2) y finales (4-5). Según las directrices de Cohen (1988), d ≥ .2, d ≥ .5 y d ≥ .8, representan tamaños del efecto pequeños, intermedios y grandes, respectivamente. El nivel de significación se fijó en p ≤ .05 a lo largo de todo el estudio. Los análisis estadísticos se realizaron con SPSS v.15 (SPSS Inc., Chicago, EE. UU.).
Resultados
La Tabla 1 muestra los valores de las áreas de histéresis y los valores iniciales de TEP, FC y O2Hb muscular para cada serie de ejercicio. Además, ilustra el modo en que el tiempo de recuperación en reposo entre series aumentó con la acumulación del esfuerzo. La Tabla 2 muestra los tamaños del efecto y las diferencias significativas en las áreas de histéresis y los valores iniciales de TEP, FC y O2Hb muscular entre las series iniciales y finales.

Tabla 1
Valores de mediana e intercuartil de las áreas de histéresis totales (suma de áreas positivas y negativas) y valores iniciales de la tasa de esfuerzo percibido (TEP), frecuencia cardiaca (FC) y oxihemoglobina muscular (O2Hb) para las cinco series consecutivas. También se incluyen la mediana y el intercuartil del tiempo de recuperación entre series.
Tabla 2
Diferencias en las áreas de histéresis entre las series iniciales y finales y los valores iniciales de tasa de esfuerzo percibido (TEP), frecuencia cardiaca (FC) y oxihemoglobina (O2Hb) muscular. La d de Cohen se representa en valores, mientras que se marcan las diferencias significativas a partir de la prueba de Wilcoxon (*p < .05).
Las áreas de FC aumentaron a lo largo de las series (χ2 = 14.16; p < .01), y las áreas de O2Hb muscular disminuyeron (χ2 = 9.84; p = .04). Las diferencias fueron significativas entre las series iniciales (1 y 2) y finales (4 y 5): FC (1-4: Z = –2.19, p = .03; 2-4: Z = –2.09, p = .04; 1-5: Z = –2.29, p = .02; 2-5: Z = –2.60, p < .01) y O2Hb muscular (1-4: Z = –2.80, p < .01; 1-5: Z = –2.19, p = .03). Tal y como se muestra en la Tabla 2, se encontraron grandes tamaños del efecto (d = 1.22 – 1.81) de las áreas de histéresis en la FC entre las series 1-4, 1-5, 2-4, 2-5, y en la O2Hb muscular entre las series 1-4 y 1-5. La TEP solo aumentó entre las series 1 y 3 y la serie 4 (Z = –1.98, p = .04; Z = –1.99, p = .04, respectivamente), con tamaños de efecto pequeños e intermedios entre las series iniciales y finales.
Todas las áreas mostraron un gran intercuartil, así como grandes diferencias entre las variables (χ2 = 59.08, p < .01). Concretamente, entre la O2Hb muscular con la FC (Z = –6.11, p < .01) y la TEP (Z = –6.00, p < .01).
Por otra parte, no hubo diferencias significativas entre los valores iniciales de series consecutivas (véanse las Tablas 1 y 2). Solo la TEP y la O2Hb muscular fueron diferentes entre las series 1 y 2 (Z = –2.35, p = .02; Z = –2.80, p < .01, respectivamente). Asimismo, se observaron tamaños del efecto pequeños e intermedios entre las series iniciales y finales de dichos valores iniciales (excepto para la FC entre las series 1 y 5).
Discusión
Este estudio, que evalúa las áreas de histéresis de la TEP, la FC y la O2Hb muscular durante series repetidas de carreras, descubrió un aumento de las áreas de TEP y FC, y una disminución de las áreas de O2Hb muscular, en las últimas series en comparación con las primeras.
Las áreas de histéresis de O2Hb muscular disminuyeron a medida que se acumulaba el esfuerzo, lo que refuerza la hipótesis del área de histéresis como posible biomarcador del estrés y la tolerancia al ejercicio (Montull et al., 2020). Ello ilustró las diferencias entre carga interna y externa (se mantuvo la VAM en todas las series) en función de la acumulación de esfuerzo. Esta diferencia también se plasmó en el aumento del tiempo de recuperación a lo largo de las series.
En contraste con las áreas de histéresis, los valores iniciales de TEP, FC y O2Hb muscular antes de las series fueron bastante similares. Estos resultados respaldan la hipótesis de que los valores cuantitativos fijos de TEP, FC y O2Hb muscular no pueden ofrecer suficiente fiabilidad como indicadores de la carga interna (Montull et al., 2020, 2022).
La propiedad de degeneración de los SAC (Edelman y Gally, 2001; Montull et al., 2020) podría explicar por qué los participantes pudieron mantener la misma velocidad a lo largo de las series a pesar de los efectos psicobiológicos de la acumulación de esfuerzo. Deben activarse diferentes sinergias psicobiológicas cuando se acumula el esfuerzo para compensar los efectos de la fatiga y cumplir el objetivo de la tarea.
Según los resultados, las sinergias psicobiológicas eran en gran medida individuales y contextuales. Las áreas de histéresis de la TEP, la FC y la O2Hb muscular variaron en gran medida a lo largo de las series, tal y como muestra su elevada dispersión estadística. Estas observaciones respaldan resultados previos que estudiaron la TEP y la FC (Montull et al., 2020) y las relaciones no proporcionales observadas entre cargas internas y externas (Impellizzeri et al., 2019).
A pesar de que el tiempo de recuperación aumentó a medida que avanzaban las series de carrera, también aumentaron las áreas de histéresis de la TEP y la FC, que pasaron de valores negativos a positivos después de la tercera serie. Esta transición indica que los valores de TEP y de recuperación de la FC fueron superiores a los del ejercicio hasta que la TEP descendió por debajo de 11. Este fenómeno ilustra la inminente inestabilidad de las sinergias psicobiológicas (Hristovski et al., 2014), que puede utilizarse como señal de alerta temprana (Scheffer et al., 2009) para ayudar a «pronosticar» el agotamiento y la desconexión de la tarea provocados por los efectos de la fatiga aguda. En consecuencia, parece posible captar dichos efectos y anticiparse a ellos utilizando el área de histéresis de las variables psicobiológicas.
La supervisión del área de histéresis también ha resultado útil para captar la eficacia de las intervenciones de entrenamiento (Martín-Guillaumes et al., 2021) y aumentar la precisión diagnóstica en cardiopatías (Zimarino et al., 2016). Así pues, estos resultados refuerzan su potencial para la supervisión de intervenciones deportivas y médicas. En el deporte, puede ayudar a gestionar de forma más precisa la recuperación durante y después del ejercicio (Montull et al., 2020). El acceso a los valores instantáneos del área de histéresis de las variables psicobiológicas durante un entrenamiento por intervalos puede permitir a los deportistas y entrenadores controlar más adecuadamente las cargas de trabajo del entrenamiento (frecuencia, duración, intensidad, recuperación). Esto va en línea con propuestas recientes orientadas a evitar tiempos de ejercicio de trabajo/descanso preestablecidos (es decir, recetas), y, en cambio, a fomentar una adaptación continua de las cargas de trabajo de entrenamiento a la evolución del estado de forma física de los deportistas y equipos (Balagué et al., 2019, 2020; Pol et al., 2020).
A pesar de su potencial, es posible que el área de histéresis de las variables fisiológicas no proporcione información precisa sobre la acumulación de esfuerzo. Las percepciones subjetivas, capaces de comprimir información múltiple de canales interoceptivos y exteroceptivos y, por tanto, de reflejar la carga de trabajo interna de forma más integrada, podrían ser un buen complemento para la supervisión (Montull et al., 2022).
Este estudio utilizó únicamente la TEP para controlar el tiempo de recuperación. Se justifica la realización de nuevos estudios que añadan otras variables como la FC para controlar el periodo de recuperación. También está justificada la realización de nuevos estudios sobre el área de histéresis de las variables psicobiológicas en poblaciones con diferentes estados de salud y de entrenamiento o forma física. Por último, este estudio subraya la importancia de desarrollar tecnología con capacidad para supervisar y evaluar la respuesta de histéresis durante el ejercicio de forma instantánea.
Conclusiones
Este estudio demostró la sensibilidad del área de histéresis de la TEP, la FC y la O2Hb muscular a la acumulación de esfuerzo durante ejercicios de carrera repetidos realizados a la VAM. El área de histéresis de las variables psicobiológicas y fisiológicas promete ser un nuevo biomarcador para controlar la fatiga aguda y evaluar los estados de salud y rendimiento.
Referencias
[1] Balagué, N., Hristovski, R., Almarcha, M., Garcia-Retortillo, S., & Ivanov, P. C. (2020). Network physiology of exercise: Vision and perspectives. Frontiers in Physiology, 11, 1607. https://doi.org/10.3389/FPHYS.2020.611550
[2] Balagué, N., Pol, R., & Guerrero, I. (2019). Science or pseudoscience of physical activity and sport? Apunts Educación Física y Deportes, 136, 129-136. http://dx.doi.org/10.5672/apunts.2014-0983.es.(2019/2).136.09
[3] Butler, D., Grood, E., Noyes, F., & Zernicke, R. (1978). Biomechanics of ligaments and tendons. Exercise and Sport Sciences Reviews, 6, 125–181.
[4] Cabasson, A., Meste, O., Bailon, R., & Laguna, P. (2012). Validation of the PR-RR hysteresis phenomenon. Computing in Cardiology, 39, 597–600.
[5] Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). New York: Routledge. https://doi.org/10.4324/9780203771587
[6] Edelman, G. M., & Gally, J. A. (2001). Degeneracy and complexity in biological systems. Proceedings of the National Academy of Sciences of the United States of America, 98(24), 13763–13768. https://doi.org/10.1073/pnas.231499798
[7] Halson, S. L. (2014). Monitoring training load to understand fatigue in athletes. Sports Medicine, 44(S2), 139–147. https://doi.org/10.1007/s40279-014-0253-z
[8] Hamaoka, T., McCully, K. K., Niwayama, M., & Chance, B. (2011). The use of muscle near-infrared spectroscopy in sport, health and medical sciences: Recent developments. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 369(1955), 4591–4604. https://doi.org/10.1098/rsta.2011.0298
[9] Hristovski, R., Balagué, N., & Schöllhorn, W. (2014). Basic notions in the science of complex systems and nonlinear dynamics. In K. Davids, R. Hristovski, D. Araújo, N. Balague, C. Button, & P. Passos (Eds.), Complex Systems in Sport (1st ed., pp. 3–17). London: Routledge. https://doi.org/10.4324/9780203134610
[10] Hristovski, R., Venskaityte, E., Vainoras, A., Balagué, N., & Vazquez, P. (2010). Constraints-controlled metastable dynamics of exercise-induced psychobiological adaptation. Medicina, 46(7), 447–453. https://doi.org/10.3390/medicina46070064
[11] Impellizzeri, F. M., Marcora, S. M., & Coutts, A. J. (2019). Internal and external training load: 15 years on. International Journal of Sports Physiology and Performance, 14(2), 270–273. https://doi.org/10.1123/ijspp.2018-0935
[12] Liu, Q., Yan, B. P., Yu, C. M., Zhang, Y. T., & Poon, C. C. Y. (2014). Attenuation of systolic blood pressure and pulse transit time hysteresis during exercise and recovery in cardiovascular patients. IEEE Transactions on Biomedical Engineering, 61(2), 346–352. https://doi.org/10.1109/TBME.2013.2286998
[13] Martín-Guillaumes, J., Montull, L., Ventura, J. L., Javierre, C., Aragonés, D., & Balagué, N. (2021). Response variability and detraining effects of standardized exercise programs. Austin Sports Medicine, 6(1):1048. https://doi.org/10.26420/austinsportsmed.2021.1048
[14] Mayergoyz, I. D. (2003). Mathematical models of hysteresis and their applications. Electromagnetism (2nd ed.). London: Academic Press.
[15] McManus, C. J., Collison, J., & Cooper, C. E. (2018). Performance comparison of the MOXY and PortaMon near-infrared spectroscopy muscle oximeters at rest and during exercise. Journal of Biomedical Optics, 23(1), 1. https://doi.org/10.1117/1.jbo.23.1.015007
[16] Medbo, J. I., & Tabata, I. (1993). Anaerobic energy release in working muscle during 30 s to 3 min of exhausting bicycling. Journal of Applied Physiology, 75(4), 1654–1660. https://doi.org/10.1152/jappl.1993.75.4.1654
[17] Montull, L., Slapšinskaitė-Dackevičienė, A., Kiely, J., Hristovski, R., & Balagué, N. (2022). Integrative proposals of sports monitoring: Subjective outperforms objective monitoring. Sports Medicine-Open, 8(41). https://doi.org/10.1186/s40798-022-00432-z
[18] Montull, L., Vázquez, P., Hristovski, R., & Balagué, N. (2020). Hysteresis behaviour of psychobiological variables during exercise. Psychology of Sport and Exercise, 48, 1–9. https://doi.org/10.1016/j.psychsport.2020.101647
[19] Nesselroade, J. R., & Molenaar, P. C. (2010). Analyzing intra-person variation: Hybridizing the ACE model with P-technique factor analysis and the idiographic filter. Behavior Genetics, 40(6), 776–783. https://doi.org/10.1007/s10519-010-9373-x
[20] Pol, R., Balagué, N., Ric, A., Torrents, C., Kiely, J., & Hristovski, R. (2020). Training or synergizing? Complex systems principles change the understanding of sport processes. Sports Medicine - Open, 6(1). https://doi.org/10.1186/s40798-020-00256-9
[21] Scheffer, M., Bascompte, J., Brock, W. A., Brovkin, V., Carpenter, S. R., Dakos, V., Held, H., Van Nes, E. H., Rietkerk, M., & Sugihara, G. (2009). Early-warning signals for critical transitions. Nature, 461(7260), 53–59. https://doi.org/10.1038/nature08227
[22] Wilmore, J. H., Costill, D. L., & Kenney, W. L. (2008). Physiology of sport and exercise. Champaign, IL: Human Kinetics.
[23] Zimarino, M., Montebello, E., Radico, F., Gallina, S., Perfetti, M., Iachini Bellisarii, F., Severi, S., Limbruno, U., Emdin, M., & De Caterina, R. (2016). ST segment/heart rate hysteresis improves the diagnostic accuracy of ECG stress test for coronary artery disease in patients with left ventricular hypertrophy. European Journal of Preventive Cardiology, 23(15), 1632–1639. https://doi.org/10.1177/2047487316655259
ISSN: 2014-0983
Recibido: 1 de julio de 2022
Aceptado: 10 de noviembre de 2022
Publicado: 1 de abril de 2023
Editado por: © Generalitat de Catalunya Departament de la Presidència Institut Nacional d’Educació Física de Catalunya (INEFC)
© Copyright Generalitat de Catalunya (INEFC). Este artículo está disponible en la url https://www.revista-apunts.com/ Este trabajo está bajo la licencia Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. Las imágenes u otro material de terceros en este artículo se incluyen en la licencia Creative Commons del artículo, a menos que se indique lo contrario en la línea de crédito. Si el material no está incluido en la licencia Creative Commons, los usuarios deberán obtener el permiso del titular de la licencia para reproducir el material. Para ver una copia de esta licencia, visite https://creativecommons.org/licenses/by-nc-nd/4.0/deed.es_ES


